

34. Наследственность и изменчивость у бактерий. Механизмы обеспечения и передачи дочерним клеткам генетической информации. Механизмы изменчивости
Наследственный материал представлен:
А. Бактериальная хромосома:
• 1 кольцевая молекула ДНК- E.coli, Shigella spp, P.aeruginosa
• 2 кольцевые хромосомы - V. cholerae, L. interrhogans, Brucellas
• Линейные хромосомы- В. burgdorfeti,Streptomyces spp
Б. Плазмиды (кольцевая и линейная форма)
Среди фенотипических признаков, сообщаемых бактериальной клетке плазмидами, можно выделить следующие:
• устойчивость к антибиотикам;
• продукцию факторов патогенности;
• способность к синтезу антибиотических веществ;
• образование колицинов;
• расщепление сложных органических веществ;
• образование ферментов рестрикции и модификации.
Репликация может происходить как и независимо от хромосомы, так и сопряжено с репликацией хромосомы.
a) Интегративные/ эписома – плазмиды, обратимо выстраивающиеся в бактериальную хромосому.
b) Трансмиссивные (конъюгативные) – способны передаваться. Трансмессивность присуща плазмидам, имеющим tra-оперон, в котором объединены гены, ответственные за перенос плазмиды- гены кодируют половые пили, которые образуют мостик с клеткой, не содержащей трансмиссивную плазмиду.
c) Мобилизируемы - Мелкие плазмиды, не несущие tra-гена, способны к передаче в присутствии в присутствии трансмис- сивных плазмид, используя их аппарат конъюгации. Процесс – мобилизация.
d) R-плазмидой – плазмиды, обеспечивающие устойчивость бактерии к антибиотикам.
e) Плазмиды, детерминирующие синтез факторов патогенности, этиологическими агентами внутрибольничных инфекций.
В. Подвижные генетические элементы.
1. Вставочные (инсерционные)последовательности — IS-элементы — это участки ДНК, способные как целое перемещаться из одного участка репликона в другой, а также между репликонами. IS-элементы содержат гены для их перемещения — транспозиции: ген, кодирующий фермент транспозазу, обеспечивающую процесс исключения IS-элемента изДНК и его интеграцию в новый локус, и ген, детерминирующий синтез репрессора, который регулирует весь процесс перемещения. Инвертированные повторы (сайты рекомбинации, окружающие вставочную последовательность) узнает фермент транспозаза, которая осуществляет одноцепочечные разрывы цепей ДНК, расположенных по обе стороны от подвижного элемента.
Подвижные элементы не являются самостоятельными репликонами, так как их репликация — составной элемент репликации ДНК репликона, в составе которого они находятся.
2. Транспозоны — это сегменты ДНК, обладающие теми же свойствами, что и IS-элементы, но имеющие в своем составе структурные гены, т.е. гены, обеспечивающие синтез молекул, обладающих специфическим биологическим свойством. (токсичностью, устойчивостью к антибиотикам).
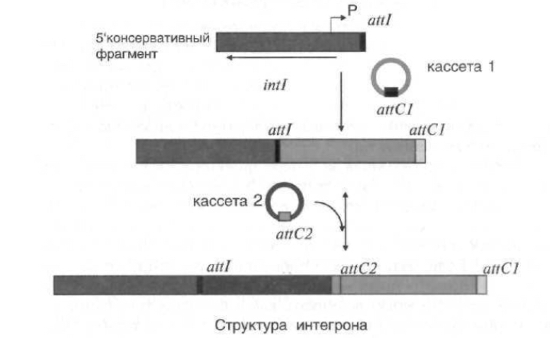
Г. Система интегронов.
- система захвата малых элементов ДНК, называемых генными кассетами, посредством сайтспецифической рекомбинации и их экспрессии.
Интегрон состоит из консервативного участка, расположенного на 5' конце, который содержит ген, кодирующий фермент интегразу, сайт рекомбинации att и промотор Р; на 3х’-конце – 1 ген антибиотикорезистентности и сайт рекомбинации.
Кассета может существовать в двух формах: линейной, когда кассета интегрирована в интегрон, и в виде маленькой кольцевой двухцепочечной ДНК.
За синтез факторов патогенности отвечают островки патогенности, которые имеют прямые повторы последовательностей ДНК или IS-элементы. Большинство островов патогенности локализовано на хромосоме бактерий (Salmonella), но также они могут находиться в составе плазмид (ShigeUa)и фаговых ДНК (V. cholerae01, 0139).
Механизмы обеспечения и передачи дочерним клеткам генетической информации.
1. Генетическая рекомбинация — это взаимодействие между двумя ДНК, обладающими различными генотипами, которое приво- дит к образованию рекомбинантной ДНК, сочетающей гены обоих родителей.
В процессе рекомбинации бактерии условно делятся на клетки-доноры, которые передают генетический материал, и клетки- реципиенты, которые воспринимают его. В клетку-реципиент проникает не вся, а только часть хромосомы клетки-донора, что приводит к формированию неполной зиготы — мерозиготы.
По молекулярному механизму генетическая рекомбинация у бактерий делится:
А. Гомологичная рекомбинация
-в процессе разрыва и воссоединения ДНК происходит обмен между участками ДНК, обладающими высокой степенью гомологии.
- процесс под контролем генов REC-системы: генов recA, B,C,D – они расплетают нити ДНК, их переориентации с образованием полухиазмы, структуры Холидея, а также разрезают структуру Холидея для завершения процесса рекомбинации.
Б. Сайтспецифическая рекомбинация
Для протекания которой необходимы строго определенные последовательности ДНК и специальный ферментативный аппарат, которые специфичны для каждого конкретного случая.
Пример:
• встраивание плазмиды в хромосому бактерий, которое происходит между идентичными IS- элементами хромосомы и плазмиды,
• интеграция ДНК фага лямбда в хромосому Е. coli.
Сайтспецифическая рекомбинация, происходящая в пределах одного репликона, участвует также в переключении активности генов.
Например, у сальмонелл следствием этого процесса являются фазовые вариации жгутикового Н-антигена.
В. Незаконная или репликативная рекомбинация
Незаконная или репликативная рекомбинация не зависит от функционирования генов гесА, В, С, D.
Пример: транспозиция подвижных генетических элементов по репликону или между репликонами, что сопровождается репликацией ДНК.
Рекомбинация осуществляется 3мя механизмами:
1. Конъюгация
- передача генетического материала от клетки-донора в клетку- репипиент путем непосредственного контакта клеток.
Необходимым условием - наличие в клетке-доноре трансмиссивной плазмиды, которые кодируют половые пили, образующие конъюгационную трубочку между клеткой-донором и клеткой-реципиентом, по которой плазмидная ДНК передается в новую клетку.
Механизм передачи плазмидной ДНК: специальный белок, кодируемый tra-опероном, узнает опре- деленную последовательность в ДНК плазмиды, вносит в эту последовательность одноцепочечный разрыв и ковалентно связывается с 5'-концом. Затем цепь ДНК, с которой связан белок, переносится в клетку-реципиент, а неразорванная комплементарная цепь остается в клетке-доноре. Клеточный аппарат синтеза ДНК достраивает одиночные цепи и в доноре, и в реципиенте до двухцепочечной структуры. Белок,свя- занный с 5'-концом перенесенной цепи, способствует замыканию плазмиды в реципиентной клетке в кольцо.
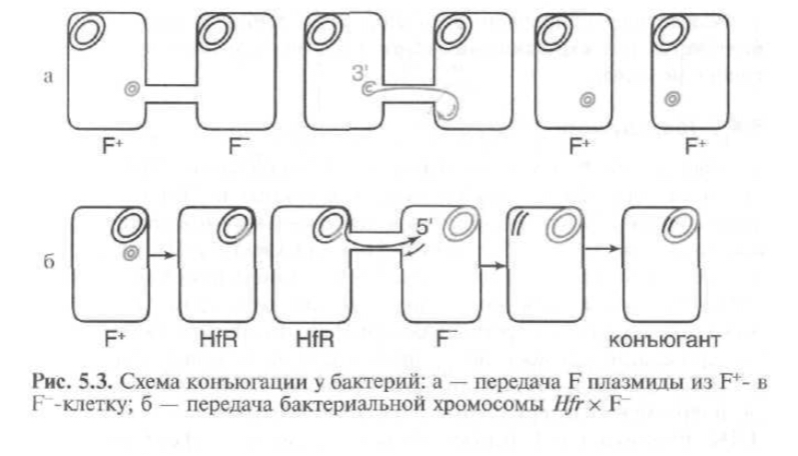
Штаммы, в которых плазмида находится в интегрированном состоянии, переносят свои хромосомные гены бесплазмидным клеткам с высокой частотой и поэтому называются Hfr.
Из-за хрупкости конъюгационного мостика половой фактор редко передается в клетку-реципиент, поэтому образовавшийся рекомбинант донорскими функциями, как правило, не обладает.
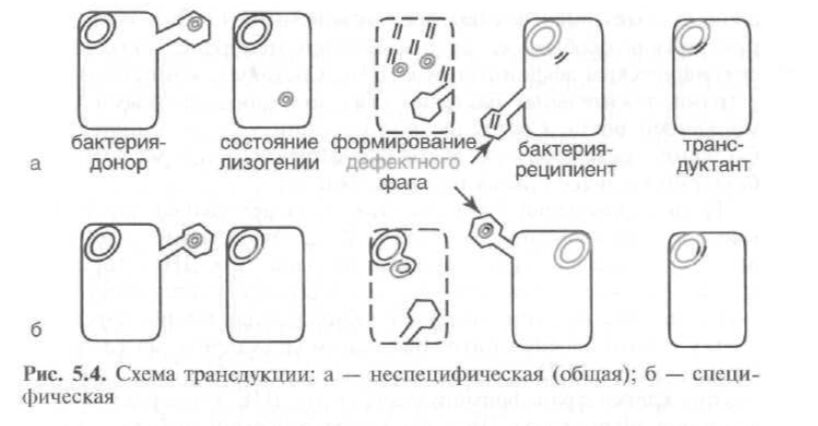
2. Трансдукция
-передача бактериальной ДНК посредством бактериофага.
В процессе репликации фага внутри бактерий фрагмент бактериальной ДНК проникает в фаговую частицу и переносится в реципиентную бактерию во время фаговой инфекции.
Типа трансдукции:
a) общая трансдукция — перенос бактериофагом сегмента любой части бактериальной хромосомы — происходит вследствие того, что в процессе фаговой инфекции бактериальная ДНК фрагментируется, и фрагмент бактериальной ДНК проникает в фаговую головку, формируя дефектную фаговую частицу и рекомбинирует гомологичной рекомбинацией с гомологичным участком хромосомы-реципиента с образованием стабильного рекомбинанта. Этим типом трансдукции обладают Р-фаги.
b) Специфическая трансдукция наблюдается в том случае, когда фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В процессе исключения ДНК- фага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы, становясь дефектным фагом. Так как большинство умеренных бактериофагов интегрирует в бактериальную хромосому в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК клетки-донора. ДНК дефектного фага рекомбинирует с ДНК клетки-реципиента сайт-специфической рекомбинацией. Рекомбинант становится меродиплоидом по привнесенному гену. В частности, бактериофаг пере- дает специфической транедукцией gal-ген у Е.coli.
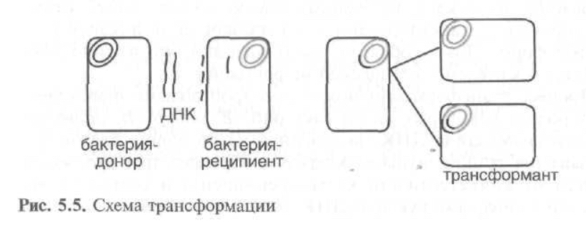
3. Трансформация
превращение бескапсульного R-штамма пневмококков (Streptococcus pneumoniae) в штамм, образующий капсулу S-формы.
Процесс может происходить самопроизвольно, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками (В. subtilis, И, influenzae, S. Pneumoniae).
Процесс трансформации зависит от компетентности, способности бактериальной клетки поглощать ДНК. У грамположительных бактерий компетентность связана с определенными фазами кривой роста, а у грамотрицательных компетентность приходится создавать искусственным путём, подвергая бактерии температурному или электрошоку.
Трансформирующей активностью обладает только двунитевая высокоспирализованная молекула ДНК, так как в то время как 1 нить ДНК проникает в клетку-реципиент, вторая подвергается деградации с высвобождением энергии, необходимой для проникновения. Рекомбинация происходит на одной нити, в результате чего образуется гетеродуготексная молекула, одна нить которой имеет генотип реципиента, а другая — рекомбинантный генотип.
Трансформация – метод генной инженерии.
Мутации:
— это изменения в последовательности отдельных нуклеотидов ДНК, которые фенотипически ведут к таким проявлениям, как изменения морфологии бактериальной клетки, возникновение потребностей в факторах роста.
• Прямая мутация – мутации, приводящие к потере функции;
• Реверсия – м., приводящие к восстановлению исходных свойств.
o Обратная/прямая реверсия – восстановление генотипа и фенотипа;
o Супрессорная – восстановление только фенотип;
По протяженности изменений повреждения ДНК различают:
Точечные – повреждение пары нуклеотидов
Протяженные/ аберрации
o выпадение нескольких пар нуклеотидов- делеция;
o Добавление нуклеотидными пар – дупликация;
o Перемещение фрагментов хромосомы – транслокации;
o Перестановки нуклеотидный пар – инверсии.
По причине возникновения:
Спонтанные – вследствие перемещения подвижных генетических элементов.
Индуцированные – под влиянием внешних факторов, мутагенез (химических, биологических -транспозоны, физических)
Замена пурина другим пурином, а пиримидина другим пиримидином называется транзицией.
Типы репарации:
• На свету – связан с деятельность фотореактивирующегося фермента, который рас- щепляет тиминовый димер.
• При темновой репарации дефектные участки цепи ДНК удаляются и образовавшаяся брешь достраивается при помощи ДНК-полимеразы на матрице сохранившейся цепи и соединяется с цепью лигазой.