

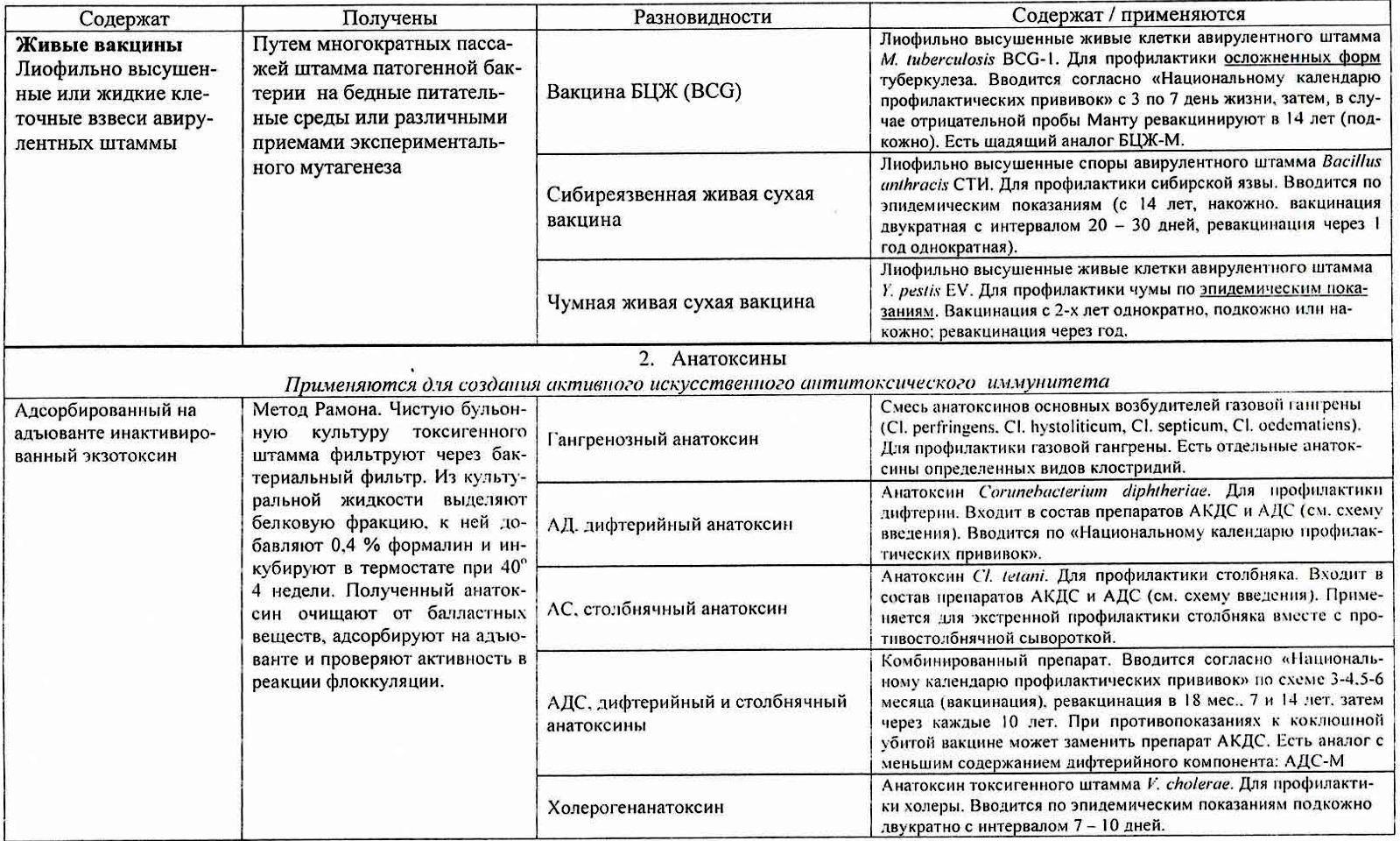
2. Анатоксины. Получение, применение. Антитоксический иммунитет
В 1923 г. Глени и Рамон обнаружили возможность превращения бактериальных экзотоксинов под влиянием формалина в нетоксичные вещества - анатоксины, обладающие антигенными свойствами. Это позволило использовать анатоксины в качестве вакцинных препаратов.
Анатоксины – препараты, полученные из бактериальных экзотоксинов, полностью лишенные своих токсических свойств, но сохранившие антигенные и иммуногенные свойства.
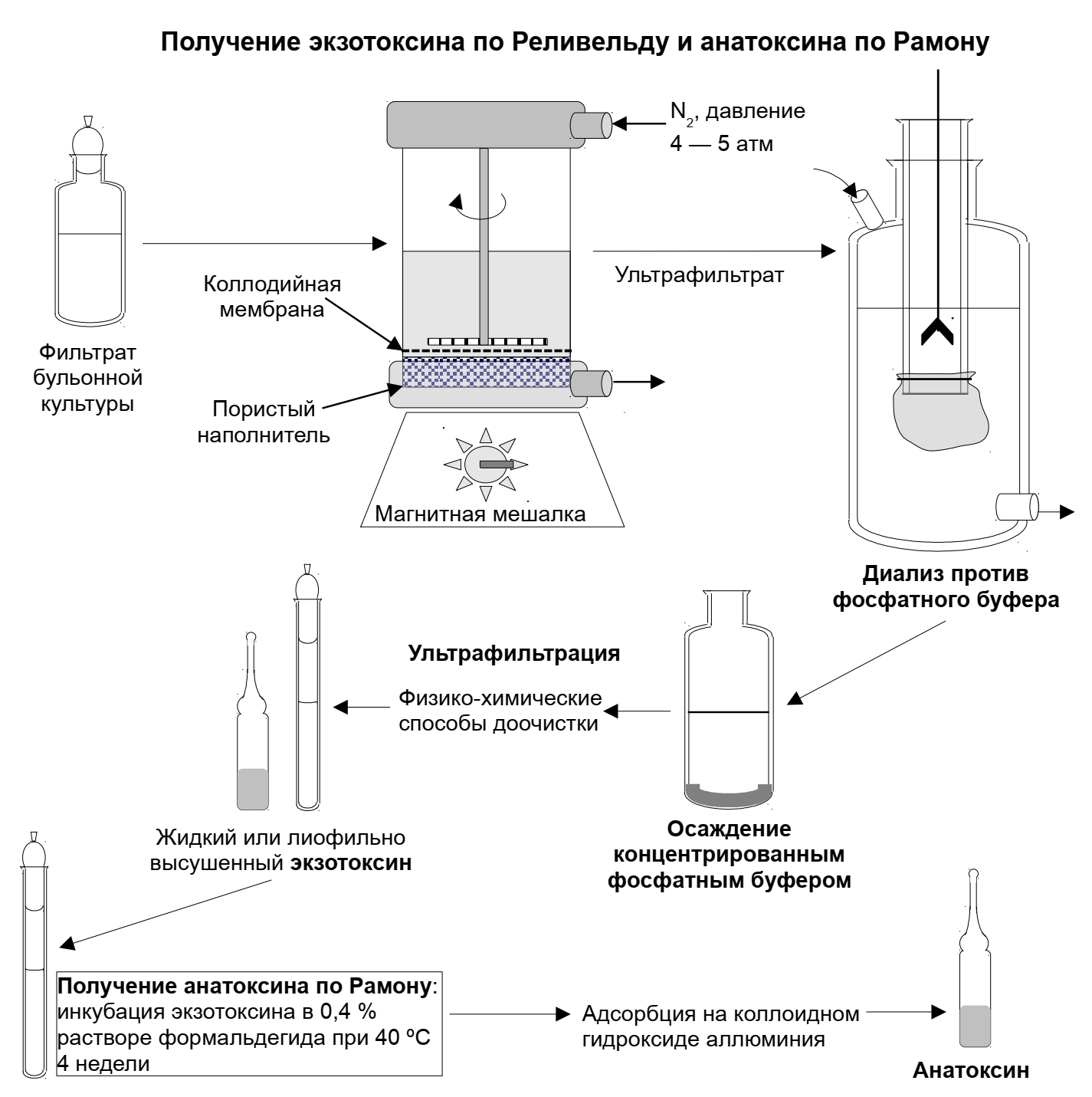
Получение: токсигенные бактерии выращивают на жидких средах, фильтруют с помощью бактериальных фильтров для удаления микробных тел, к фильтрату добавляют 0,4% формалина и выдерживают в термостате при 30-40t на 4 недели до полного исчезновения токсических свойств, проверяют на стерильность, токсигенность и иммуногенность. Эти препараты называются нативными анатоксинам, в настоящее время почти не используются, т. к. содержат большое количество балластных веществ, неблагоприятно влияющих на организм. Анатоксины подвергают физической и химической очистке, адсорбируют на адъювантах. Такие препараты называются адсорбированными высокоочищенными концентрированными анатоксинами.
Титрование анатоксинов в реакции фолликуляции производят по стандартной фолликулирующей антитоксической сыворотке, в которой известно количество антитоксических единиц. 1 антигенная единица анатоксина обозначается Lf, это то количество анатоксина, которое вступает в реакцию флокуляции с 1 единицей дифтерийного анатоксина.
Применение: Анатоксины применяются для профилактики и реже, для лечения токсинемических инфекций (дифтерия, газовая гангрена, ботулизм, столбняк). Так же анатоксины прим
еняются для получения антитоксических сывороток путем гипериммунизации животных.
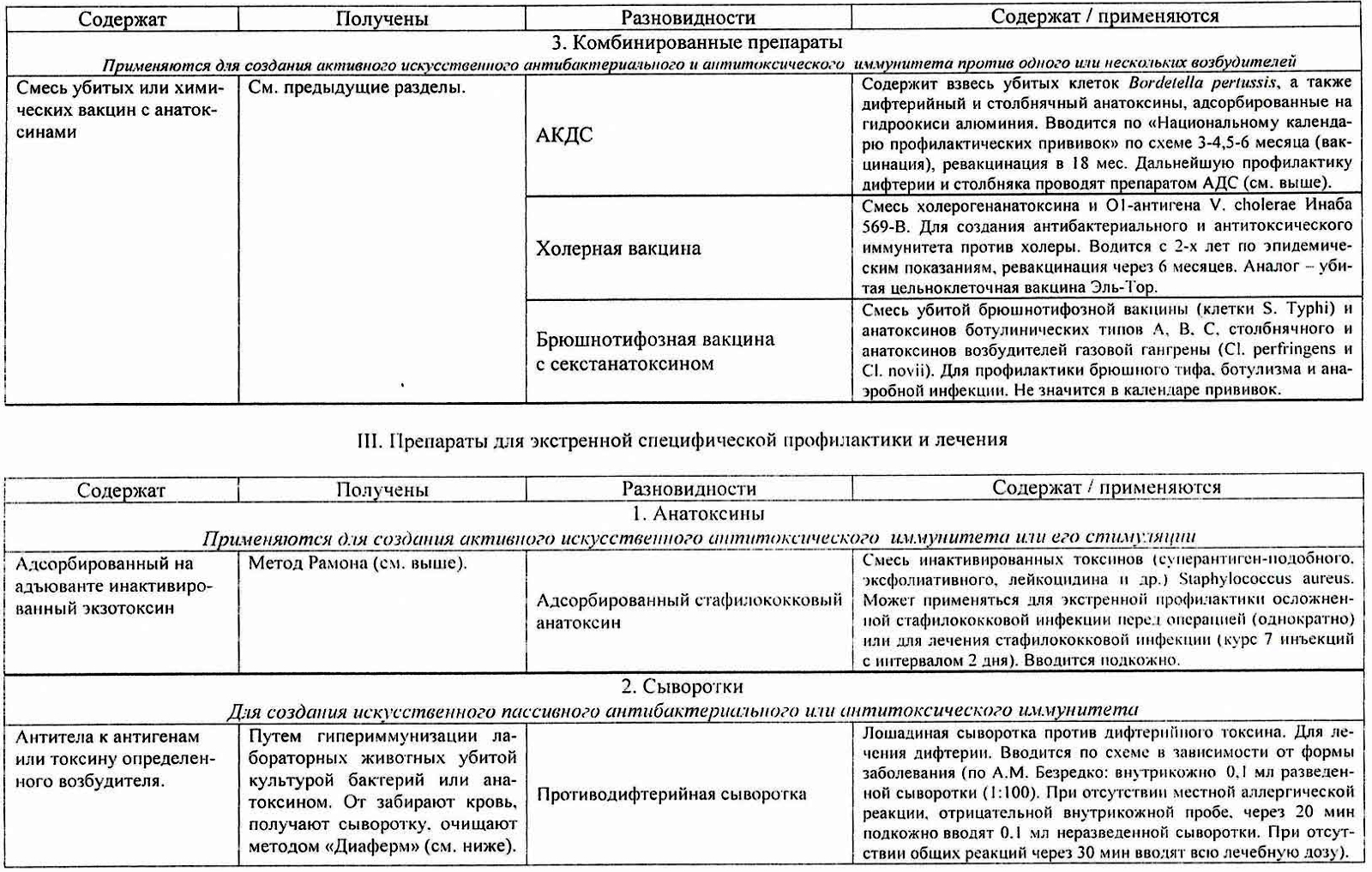
Примеры препаратов: АКДС, АДС, адсорбированный стафилококковый анатоксин, ботулинистический анатоксин, анатоксины из экзотоксинов возбудителей газовых инфекций.
Цель применения анатоксинов — индукция иммунных реакций, направленных на нейтрализацию токсинов; в результате иммунизации синтезируются нейтрализующие AT (антитоксины). Обычный источник токсинов — промышленно культивируемые естественные штаммы-продуценты (например, возбудители дифтерии, ботулизма, столбняка). Полученные токсины инактивируют термической обработкой либо формалином, в результате чего образуются анатоксины (токсоиды), лишённые токсических свойств, но сохранившие иммуногенность. Анатоксины очищают, концентрируют и для усиления иммуно-генных свойств адсорбируют на адъюванте (обычно, гидрооксид алюминия). Адсорбция анатоксинов значительно повышает их иммуногенную активность. С одной стороны, образуется депо препарата в месте его введения с постепенным поступлением в кровоток, с другой — действие адъюванта стимулирует развитие иммунного ответа, в том числе и в регионарных лимфатических узлах. Анатоксины выпускают в форме моно- (дифтерийный, столбнячный, стафилококковый) и ассоциированных (дифтерийно-столбнячный, ботулинический трианатоксин) препаратов.
Дополнительно: Научная электронная библиотека (monographies.ru)
Антитоксический иммунитет – отдельные бактерии в организме выделяют токсины, на которые вырабатываются антитела, нейтрализующие токсины в организме животного.
Иммунитет антитоксический – невосприимчивость организма к инфекционным болезням, возбудители которых продуцируют экзотоксины. Иммунитет антитоксический достигается активной иммунизацией, введением в организм анатоксина, вызывающего синтез антитоксинов, антитоксических сывороток или пассивной иммунизацией. Содержание антитоксинов в антитоксических сыворотках выражается в антитоксических единицах (АЕ). О напряженности антитоксического иммунитета можно судить по содержанию антитоксинов в сыворотке крови. Антитоксический и антимикробный иммунитет. Различают антитоксический иммунитет, направленный на нейтрализацию микробных ядов, и антибактерийный иммунитет, направленный на уничтожение самих микробных тел. В наиболее чистом виде антитоксический иммунитет проявляется при токсических инфекциях (дифтерия, столбняк, ботулизм, газовая гангрена и др.).
В механизме антитоксического иммунитета имеет значение не только наличный титр антитоксинов в крови иммунного человека или животного, но и способность организма к их выработке.
Конспект преподавателя:
Генетические рекомбинации — это взаимодействие между двумя молекулами ДНК, обладающими различными генотипами, которое приводит к образованию рекомбинантной ДНК, сочетающей гены обоих молекул. Особенности рекомбинации у бактерий определяет отсутствие полового размножения и мейоза, в процессе которых у высших организмов происходят рекомбинация, а также гаплоидный набор генов.
В процессе рекомбинации бактерии условно делятся на клетки-доноры, которые передают генетический материал, и клетки-реципиенты, которые воспринимают его. В клетку-реципиент проникает не вся, а только часть хромосомы клетки-донора, что приводит к формированию неполной зиготы — мерозиготы. В результате рекомбинации в мерозиготе образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента,
По молекулярному механизму генетическая рекомбинация у бактерий делится на гомологичную, сайтспецифическую и незаконную.
При гомологичной рекомбинации в процессе разрыва и воссоединения ДНК происходит обмен между участками ДНК, обладающими высокой степенью гомологии. Процесс гомологичной рекомбинации находится под контролем генов, объединенных в RЕС -систему.
Сайтспецифическая рекомбинация - этот тип рекомбинации не зависит от функционирования RЕС –генов, не требует протяжных участков гомологии ДНК, но для протекания которой необходимы строго определенные последовательности ДНК и специальный ферментативный аппарат, специфичный для каждого конкретного случая. Примером этого типа рекомбинации является встраивание плазмиды в хромосому бактерий, которое происходит между идентичными IS-элементами хромосомы и плазмиды, интегра-ция ДНК фага лямбда в хромосому Е. coli.
Незаконная или репликативная рекомбинация - также не зависит от функционирования генов RЕС. Примером ее является транспозиция подвиж-ных генетических элементов по репликону или между репликонами, при этом, транспозиция подвижного генетического элемента сопровождается репликацией ДНК.
Генетические рекомбинации могут осуществляться путем трансформации, трансдукции, конъюгации, слияния протопластов.
Трансформация - процесс трансформации может самопроизвольно происходить в природе у некоторых видов бактерий, В. subtilis, И, influenzae, S. pneumoniae, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками. Процесс трансформации зависит от компетентности клетки-реципиента и состояния донорской трансформирующей ДНК. Компетентность — это способность бактериальной клетки поглощать ДНК.
Она зависит от присутствия особых белков в клеточной мембране, обладающих специфическим аффинитетом к ДНК. Состояние компетентности у грам положительных бактерий связано с определенными фазами кривой роста. Состояние компетенции у грамотрицательных бактерий приходится созда-вать искусственным путем, подвергая бактерии температурному или элек-трошоку.
Трансформирующей активностью обладает только двунитевая высокос-пирализованная молекула ДНК. Это связано с тем, что в клетку-реципиент проникает только одна нить ДНК, тогда как другая — на клеточной мембране — подвергается деградации с высвобождением энергии, которая необходима для проникновения в клетку сохранившейся нити. Высокая молекулярная масса трансформирующей ДНК увеличивает шанс рекомбинации, так как внутри клетки трансформирующая нить ДНК подвергается воздействию эндонуклеаз. Интеграция с хромосомой требует наличия гомологичных с ней участков у трансформирующей ДНК. Рекомбинация происходит на одной нити, в результате чего одна нить имеет генотип реципиента, а другая — рекомбинантный генотип. Рекомбинантные трансформанты формируются только после цикла репликации.
Конъюгация – осуществляется при непосредственном контакте клеток. Контролируется tra- (transfer) опероном. Главную роль играют конъюгативные F- плазмиды, которые кодируют половые пили, образующие конъюгационный мостик (трубочку) между клеткой-донором и клеткой-реципиентом, по которой плазмидная ДНК передается в клетку-реципиент. Если F-фактор находится в клетке-доноре в автономном состоянии, то в результате взаимодействия клетка-реципиент приобретает донорские свойства. Если F-фактор или другая трансмиссивная плазмида встраиваются в хромосому клетки-реципиента, то плазмида и хромосома начинают функционировать в виде единого трансмиссивного репликона, что делает возможным перенос бактериальных генов в бесплазмидную клетку-реципиент.
Трансдукция – это передачу бактериальной ДНК посредством бакте-риофага. В процессе репликации фага внутри бактерий фрагмент бактериальной ДНК проникает в фаговую частицу и переносится в реципиентную бактерию во время фаговой инфекции. Существует два типа трансдукции: общая трансдукция — перенос бактериофагом сегмента любой части бактериальной хромосомы — происходит вследствие того, что в процессе фаговой инфекции бактериальная ДНК фрагментируется, и фрагмент бактериальной ДНК того же размера, что и фаговая ДНК, проникает в фаговую головку, формируя дефектную фаговую частицу – умеренный бактериофаг. Этот процесс происходит с частотой приблизительно 1 на 1000 фаговых частиц. При инфицировании клетки-реципиента дефектной фаговой частицей ДНК клетки-донора впрыскивается в нее и рекомбинирует путем гомологичной рекомбинацией с гомологичным участком хромосомы-реципиента с образованием стабильного рекомбинанта.
Специфическая трансдукция наблюдается в том случае, когда фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В обратном процессе дезинтеграции ДНК-фага из бактериальной хромосомы случайным образом захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы, бактериофаг при этом становится дефектным, т.е. умеренным. В дальнейшем, при следующем взаимодействии такого умеренного бактериофага с бактериальной клеткой происходит его интегрирация в хромосому в строго специфических участках ДНК клетки-реципиента путем сайт-специфической рекомбинацией и таким образом в структуру ДНК клетки-реципиента включается и фрагмент ДНК клетки-донора.