Достоинства метода
· Простой метод, доступный для практических лабораторий
· Количественная характеристика резистентности
Недостатки метода
· Высокая стоимость
· Большой расход питательных сред (на стандартную чашку Петри можно поместить только один тест)
Доп инфа:
Определение антибиотика в крови, моче и других жидкостях организма человека.
В штатив устанавливают два ряда пробирок. В одном из них готовят разведения эталонного антибиотика, в другом — исследуемой жидкости.
Затем в каждую пробирку вносят взвесь тест-бактерий, приготовленную в среде Гисса с глюкозой.
При определении в исследуемой жидкости пенициллина, тетрациклинов, эритромицина в качестве тест-бактерий используют стандартный штамм S. aureus, а при определении стрептомицина — Е. coli.
После инкубирования посевов при 37 °С в течение 18—20 ч отмечают результаты опыта по помутнению среды и ее окрашиванию индикатором вследствие расщепления глюкозы тест-бактериями.
Концентрация антибиотика определяется умножением наибольшего разведения исследуемой жидкости, задерживающей рост тест-бактерий, на минимальную концентрацию эталонного антибиотика, задерживающего рост тех же тестбактерий.
Например, если максимальное разведение исследуемой жидкости, задерживающее рост тест-бактерий, равно 1 :1024, а минимальная концентрация эталонного антибиотика, задерживающего рост тех же тест-бактерий, 0,313 мкг/мл, то произведение 1024- 0,313=320 мкг/мл составляет концентрацию антибиотика в 1 мл.
Определение способности S. aureus продуцировать бета-лактамазу.
В колбу с 0,5 мл суточной бульонной культуры стандартного штамма стафилококка, чувствительного к пенициллину, вносят 20 мл расплавленного и охлажденного до 45 °С питательного агара, перемешивают и выливают в чашку Петри. После застывания агара в центр чашки на поверхность среды помещают диск, содержащий пенициллин. По радиусам диска петлей засевают исследуемые культуры. Посевы инкубируют при 37 °С до следующего дня, после чего отмечают результаты опыта. О способности исследуемых бактерий продуцировать бета-лакта-мазу судят по наличию роста стандартного штамма стафилококка вокруг той или другой исследуемой культуры (вокруг диска).
25. Механизм фагоцитоза. Выживание микобактерий туберкулеза в фагоците. Туберкуломы.
Общая информация по учебнику Зверева.
Фагоцитоз – процесс поглощения клеткой частиц или крупных макромолекулярных комплексов, складывающийся из нескольких этапов:
Активация и хемотаксис – целенаправленное движение клеток к объекту фагоцитоза в сторону повышающейся концентрации хемоаттрактантов (хемокины, компоненты комплемента, продукты деградации тканей);
Адгезия частиц к поверхности фагоцита. В адгезии важную роль играют Toll-подобные рецепторы, а также рецепторы к Fc-фрагменту иммуноглобулина и C3в-компоненту комплемента (иммунный фагоцитоз).
Поглощение частиц – погружение в цитоплазму и образование фагосомы(вакуоли).
Внутриклеточный киллинг и переваривание. Фагосома сливается с лизосомой→фаголизосома, в которой под действием ферментом происходит гибель бактерии (кислороднезависимая система бактерицидности). Одновременно в клетке усиливается потребление кислорода и глюкозы, развивается респираторный взры (образуются токсичные продукты кислорода, н-р, перекись водорода и супероксиданион) – кислородзависимая система бактерицидности.
*Фагоциты – это макрофаги (моноциты и тканевые макрофаги) и микрофаги (нейтрофилы).
Касаемо туберкулеза и фагоцитоза:
Фагосома→фаголизосома→ в результате процессирования МБТ в фаголизосоме фрагменты микобактерии презентуются на поверхность макрофага. Исходы попадания МБТ в фаголизосому:
• завершенный фагоцитоз – уничтожение МБТ макрофагом;
• сохранение (выживание) ее в макрофаге.
• незавершенный фагоцитоз - внутриклеточное размножение МБТ → гибель альвеолярного макрофага→повторный фагоцитоз;
При незавершенном фагоцитозе МБТ происходит апоптоз, тем самым обеспечивая важную связь с адаптивным иммунитетом, поскольку апоптозные везикулы содержат антигены МБТ, которые захватываются ДК (дендритные клетки). Последние презентуют эти антигены незрелым Т-клеткам, стимулируя их активацию.
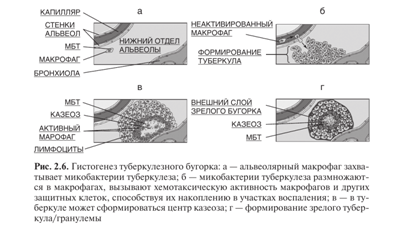
Одновременно макрофаги выделяют в окружающую среду противовоспалительные цитокины и интерлейкины, притягивающие в очаг инфекции Т-лимфоциты. Т-лимфоциты считывают информацию об антигенах с мембраны макрофага и превращаются в клетки памяти. Таким образом, активированные Т-лимфоциты продуцируют эффекторные цитокины, гамма-интерферон (IFN-y) и фактор некроза опухоли альфа (TNF-a), которые привлекают из капилляров в очаг воспаления большое количество различных клеток: макрофагов, лимфоцитов, гранулоцитов и др. Эти клетки участвуют в формировании гранулемы и тем самым ограничивают распространение МБТ в организме. На месте воспаления происходит формирование специфических бугорков — гранулем/туберкулем— как проявление локального туберкулезного воспаления в участке своего первоначального внедрения.
Подробнее про сохранение в макрофаге
Бактерицидной активности. Проникновение МБТ не сопровождается полноценной активацией макрофагов (из-за особенности клеточной стенки МБТ)→фагоцитоз не сопровождается образованием перекисных радикалов и носит неагрессивный характер. Далее проникнув внутрь, микобактерии включают механизмы, препятствующие образованию фаголизосом (продукция аммония, синтез сульфолипидов).
Если же фаголизосома образовалась, то МБТ начинают подавлять кислородозависимую и кислородонезависимую киллерную активность макрофагов целым рядом механизмов:
Синтез ферментов с каталазной и пероксидазной активностью;
Продолжающийся синтез аммония и сульфолипидов.
Используя железосодержащие соединения макрофагов для своих ферментных систем, МБТ блокируют иммуноспецифичемкие функции макрофагов (снижение антигенпредставляющей ункции, ослабление чувствительности к активирующим сигналам Т-лимфоцитов). Поражаются и митохондрии макрофагов (миколовыми арабинолипидными кислотами), что в конечном счете может привести к апоптозу.
26 . Механизмы возникновения и распространения устойчивости бактерий к антибактериальным препаратам. Природная и приобретенная устойчивость к антибактериальным препаратам.
Антибиотикорезистентность – феномен устойчивости микробных штаммов к действию одного или нескольких антимикробных препаратов (АМП). В микробиологии – способность штамма выживать при более высоких концентрациях АМП, чем большинство штаммов этого вида. Антибиотикорезистентость – пример адаптации бактерий к условиям внешней среды.
Различия внебольничных и внутрибольничных (госпитальных) инфекций – во внебольничных инфекциях: этиологический агент специфичен для отдельной нозологической формы (определенной болезни), уровень АБ-резистентности можно предсказать, возможно разработать универсальные рекомендации для терапии. Внутрибольничные инфекции – самые устойчивые, выраженной специфичности болезни нет, универсальные рекомендации не разработать.
Различают природную (видовую, первичную, естественную) и приобретенную устойчивость.
Видовая устойчивость
· является постоянным видовым признаком
· Детерминируется хромосомой
· Отсутствие мишени для АМП или недоступность мишени из-за инзкой проницаемости ЦПМ или ферментативной инактивации АПМ
· При её наличии у бактерий препараты не эффективны, она легко прогнозируется
Приобретенная устойчивость
· свойство отдельных штаммов бактерий сохранять жизнеспособность при концентрациях антибиотиков, которые подавляют основную часть микробной популяции
· детерминируется хромосомой (мутации, транспозоны) или R-плазмидами
· эффективность препарата не всегда снижается
Генетические механизмы АБ-резистентности
· Мутации
· Рекомбинации (перераспределение генов с созданием новых комбинаций)
· Репликативные единицы
o Конъюгативные транспозоны – мобильные генетические элементы
o Конъюгативные R-плазмиды – способные передаваться в другие клетки
· Нерепликативные едицины
o Транспозоны
o Генные кассеты – автономные, нереплицирующиеся, замкнутые в кольцо элементы.
o Интегрон – система захвата и экспрессии генов, например, в генной кассете
Биохимические механизмы развития устойчивости к химиотерапевтическим препаратам.
1. Выработка ферментов, инактивирующих АБ
2. Снижение проницаемости ЦМП бактерии для АБ
3. Структурные изменения мишени
4. Альтернативные пути метаболизма
5. Эффлюкс – актиный выброс АБ из клетки
Диагностика резистентности:
o Диско-диффузионный метод
o Е-тест
o Метод серийных разведений
27. Механизмы обмена генетической информацией у бактерий
Рекомбинация – процесс генетического обмена между донором и реципиентом, в результате которого является включение фрагментов нуклеиновой кислоты донора в хромосому реципиента путём кроссинговера.
Передача генетического материала (хромосомных генов) от одних бактерий к другим происходит путем:
1) трансформации,
2) трансдукции
3) конъюгации,
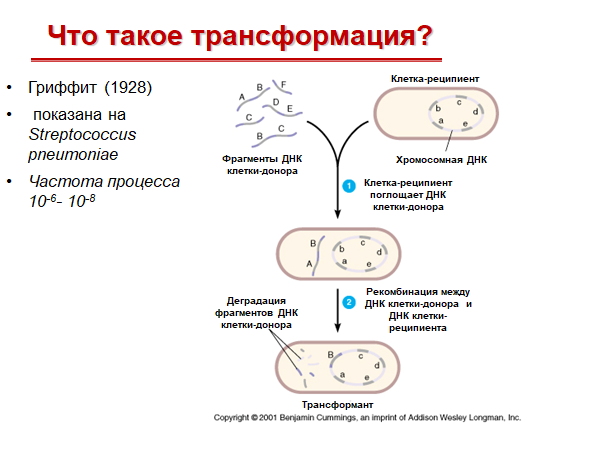
Трансформация – фрагмент нативной ДНК клетки донора, оказавшийся в окружающей среде, поглощается реципиентом (зачастую после смерти донора)
В 1944 г. О. Эвери, К. Мак-Леод и К. Мак-Карти установили, что активным началом, содержащимся в экстракте убитых пневмококков, является ДНК, которая определяет его генетические свойства и является носителем генетической информации. Феномен трансформации воспроизводится в опытах с разными патогенными и непатогенными бактериями: стрептококками, менингококками и др. С донорной ДНК в реципиентную клетку обычно передается только один ген. Это связано с протяженностью трансформирующего фрагмента ДНК, который может проникнуть в реципиентную клетку. Обычно он не превышает 1/100 длины бактериальной хромосомы, т.е. включает один или несколько сцепленных генов. Эффективно трансформация происходит в опытах с бактериями одного и того же вида, имеющих разный генотип.
Процесс трансформации бактерий можно подразделить на несколько фаз:
1. адсорбция ДНК-донора на клетке-реципиенте;
2. проникновение ДНК внутрь клетки-реципиента;
3. соединение ДНК с гомологичным участком хромосомы реципиента с последующей рекомбинацией.
После проникновения внутрь клетки трансформирующая ДНК деспирализуется. Затем происходит физическое включение любой из двух нитей ДНК донора в геном реципиента.
Эффективность спаривания трансформирующей ДНК с соответствующим участком хромосомы реципиента зависит от степени гомологичности ДНК донора и реципиента. Чем выше гомологичность, тем эффективнее спаривание, что определяет конечный результат трансформации, т.е. количество формирующихся рекомбинантов (трансформантов). Отсюда ясно, почему межвидовая трансформация происходит гораздо реже, чем внутривидовая
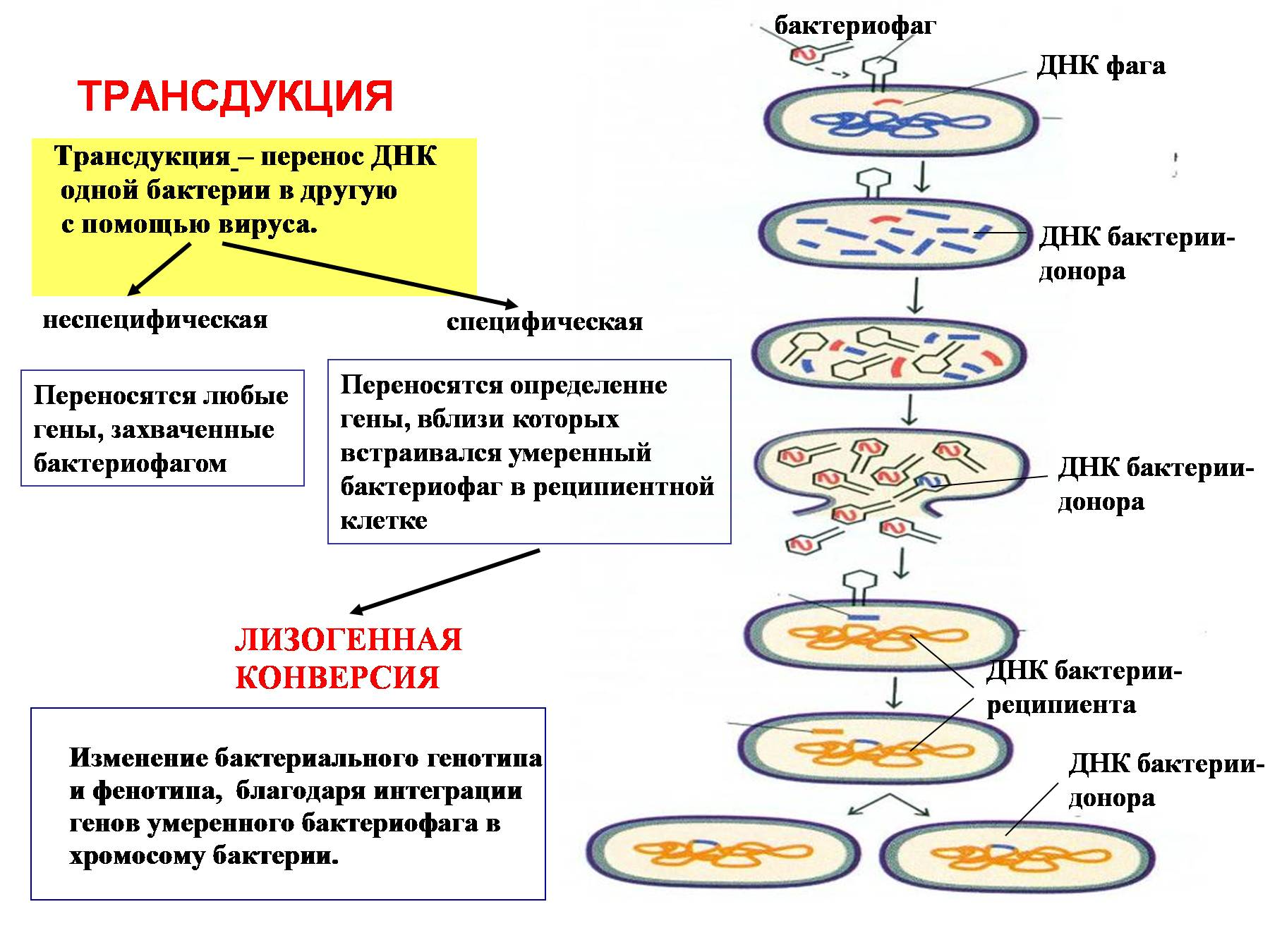
Трансдукция – перенос генетической информации от донора к реципиенту при помощи умеренного бактериофага. Есть специфическая (когда кусок профага образует новую ДНК для следующего поколения фагов) и неспецифическая/общая (когда при формировании нового поколения фагов в капсулу попадает не фаговая генетическая информация, а бактериальная)
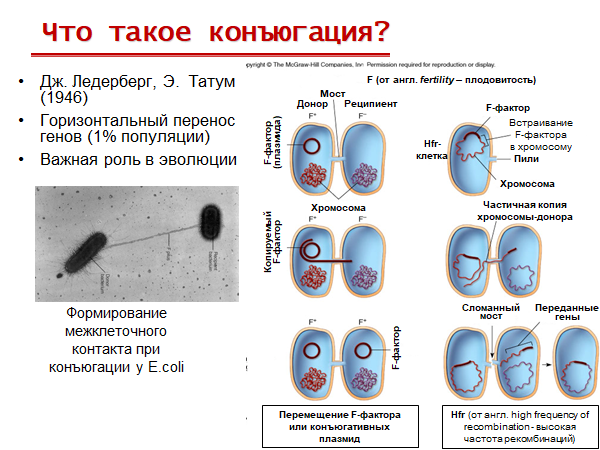
Конъюгация – «половой процесс» у бактерий, при котором в результате физического контакта через sex-пили может происходить передача ДНК от донора к реципиенту и рекомбинация. Для неё нужна F-плазмида (на ней кодируется специальный белок, который может разрезать 1 из 2 цепей основной ДНК и перетащить её по половым пили к реципиенту, тогда оставшаяся цепь восстановит ту, что у неё забрали, а та, которая ушла реципиенту, может стать плазмидой, также восстановившись). F-плазмида переходит к реципиенту вместе с транспортируемы участком по наследству.
Этапы конъюгации:
1. Первым этапом конъюгации является прикрепление клетки-донора к реципиентной клетке с помощью половых ворсинок (sex pili).
2. Затем между обеими клетками образуется конъюгационный мостик, через который из клетки-донора в клетку-реципиент могут передаваться F-фактор и другие плазмиды, находящиеся в цитоплазме бактерии-донора в автономном состоянии.
Для переноса бактериальной хромосомы необходим разрыв одной из цепей ДНК, который происходит в месте включения F-плазмиды при участии эндонуклеазы.
3. Проксимальный конец ДНК через конъюгационный мостик проникает в клетку-реципиент и сразу же достраивается до двунитевой структуры. Оставшаяся в клетке донора нить ДНК является матрицей для синтеза второй нити. Следовательно, при конъюгации передается только одна нить ДНК-донора, а вторая, оставшаяся комплементарная, цепь достраивается в реципиентной клетке.