

76. Дыхание: этапы; механизмы вдоха и выдоха. Значение дыхательных мышц в обеспечении внешнего дыхания. Эластическая тяга легких, факторы, ее определяющие. Сурфактант, его значение.
Дыхание- это физиологический процесс, обеспечивающий обмен кислородом и углекислым газом между окружающей средой и организмом.
Стадии дыхания:
Внешнее дыхание - обмен кислородом и углекислым газом между окружающей средой и кровью легочных капилляров
Легочная вентиляция – обмен между внешней средой и альвеолами – конвекция (на далекие расстояния, процесс активный с затратой энергии)
Легочная диффузия – газообмен между кровью организма и газовой смесью в легких (на короткие расстояния, менее чем на 0,1 м, процесс пассивный.)
Транспорт газов кровью – путём конвекции;
Тканевая диффузия – обмен кислородом и углекислым газом
Тканевое дыхание – потребление кислорода и выделение углекислого газа
Механизм вдоха:
Возбуждение -> сокращение респираторных мышц;
Сокращение диафрагмы, увеличение вертикального объема грудной клетки (ДО70%);
Диафрагма опускается на 1 см;
Объём лёгких увеличивается;
Внутрилегочное давление уменьшается до -1 мм.рт.ст. – меньше атмосферного;
Воздух по градиенту поступает в альвеолы.
Механизм выдоха:
Расслабление испираторных мышц;
Диафрагма поднимается;
Объём грудной клетки уменьшается;
Объём лёгких уменьшается;
Внутрилегочное давление повышается до +1мм.рт.ст – больше атмосферного.;
Воздух по градиенту выходит из лёгких.
Дыхательные мышцы:
Инспираторные (обеспечивают увеличение объема грудной клетки и легких -> вдох):
Основные: диафрагма, наружные межреберные и межхрящевые;
Вспомогательные: большие и малые грудные, лестничные, ГКС, зубчатые.
Экспираторные (обеспечивают уменьшение грудной клетки и легких -> выдох): внутренние межреберные и мышцы передней брюшной стенки.
Эластическая тяга легких — сила, с которой ткань легкого противодействует атмосферному давлению и обеспечивает спадение альвеол (обусловлена наличием в стенке альвеол большого количества эластических волокон и поверхностным натяжением пленки жидкости, покрывающей внутреннюю поверхность альвеол), т. е. это сила, с которой легкие стремятся сжаться.
Эластическая тяга легких обусловлена 3 факторами:
1.Поверхностным натяжением пленки жидкости, покрывающей внутреннюю поверхность альвеол – сурфактанта;
2.Упругостью ткани стенок альвеол (содержат эластические волокна).
3.Тонусом бронхиальных мышц.
Важным фактором, влияющим на эластические свойства и растяжимость легких, является поверхностное натяжение жидкости в альвеолах. Спадению альвеол препятствует сурфактант, выстилающий внутреннюю поверхность альвеол, препятствующий их спадению, а также выходу жидкости на поверхность альвеол из плазмы капилляров легкого. На любой поверхности раздела между воздухом и жидкостью действуют силы межмолекулярного сцепления, стремящиеся уменьшить величину этой поверхности (силы поверхностного натяжения). Под влиянием этих сил альвеолы стремятся сократиться. Поверхностное натяжение альвеол в 10 раз меньше теоретически рассчитанного натяжения для соответствующей водной поверхности.
Силы поверхностного натяжения создают 2/3 эластической тяги легких. Если заполнить легкие водой, поверхностное натяжение увеличится в 5–8 раз. Следовательно, в состав жидкости, покрывающей внутреннюю поверхность альвеол, входит вещество, снижающее поверхностное натяжение, — сурфактант.
Он представляет собой мономолекулярный слой строго ориентированных молекул поверхностно-активных веществ — липопротеин (90 % составляют липиды, преимущественно фосфолипиды, дипальмитоилфосфатидилхолин — 45 %, фосфатидилхолин — 25 %, фосфатидилглицерол — 5 %, остальные фосфолипиды — 5 %, другие липиды (холестерин, триглицериды, ненасыщенные жирные кислоты, сфигномиелин) — 10 %). Оставшиеся 10 % приходятся на долю белковой фракции, которая представлена белками-апопротеинами.
Сурфактант синтезируется альвеолоцитами 2-го типа, отработанные сурфактанты поглощаются альвеолярными макрофагами и альвеолоцитами 3-го типа. Слой сурфактанта имеет толщину 20–100 нм. Когда легкие растягиваются, пленка сурфактанта становится менее плотной и в меньшей степени уменьшает поверхностное натяжение. При уменьшении размеров альвеол поверхностное натяжение, напротив, уменьшается, что препятствует их спаданию. Нарушение образования или действия сурфактанта сопровождается спадением большого количества альвеол, в результате чего возникает ателектаз (ateles (греч.) — неполный, ektasis (греч.) — расширение) — состояние, при котором обширные участки легких не вентилируются.
Синтез и замена поверхностно-активного вещества — сурфактанта происходит довольно быстро, поэтому нарушение кровотока в легких, воспаление и отеки, курение, острая кислородная недостаточность (гипоксия) или избыток кислорода (гипероксия), а также различные токсические вещества, в том числе некоторые фармакологические препараты (жирорастворимые анестетики), могут снизить его запасы и увеличить поверхностное натяжение жидкости в альвеолах. Все это ведет к их ателектазу, или спадению. В профилактике и лечении ателектазов определенное значение имеют аэрозольные ингаляции лекарственных средств, содержащих фосфолипидный компонент, например лецитин, который способствует восстановлению сурфактанта.
Функции сурфактанта:
обеспечивает расправление легких при первом вдохе новорожденного,
в 10 раз уменьшает силу поверхностного натяжения,
стабилизирует размеры альвеол,
способствует переключению дыхания с одних альвеол на другие,
уменьшает энергозатраты на дыхание,
регулирует водный баланс (влияет на интенсивность испарения воды с альвеолярной поверхности),
способствует сохранению сухой поверхности альвеол,
облегчает диффузию кислорода из альвеол в кровь, обладает бактериостатическим действием (опсонизация бактерий),
оказывает противоотечное, антиокислительное действие,
способствует активизации фагоцитоза альвеолярными макрофагами.
Эластическая тяга грудной клетки создается за счет эластичности межреберных хрящей, мышц, париетальной плевры, структур соединительной ткани, способных сжиматься и расширяться. Благодаря эластической тяге давление в плевральной полости (Рpl) ниже давления в альвеолах (Ра) на величину, ей создаваемую. Поэтому транспульмональное давление (Рt), представляющее собой разность Ра-Рpl , стремится расправить легкие. При вдохе, когда Рpl, как указывалось выше, становится более отрицательным, Рt возрастает. Вследствие этого объем легких увеличивается, а давление в них уменьшается. Когда Ра становистся ниже атмосферного, воздух устремляется в легкие.
Дополнительно:
Неэластическая тяга- сила трения внутри воздушной струи и между потоком воздуха и стенкой дыхательных путей и аэродинамическим сопротивлением.
Величина аэродинамического сопротивления зависит от :
Трения между молекулами газовых смесей;
Вязкости газов;
Длины воздухоносных путей.
77. Газообмен в легких. Состав атмосферного и альвеолярного воздуха. Диффузия газов в легких. Факторы, определяющие объем диффузии. Напряжение газов в артериальной и венозной крови, тканях. Кислородная емкость крови. Коэффициент утилизации кислорода.
Альвеолярная вентиляция (МВЛ) характеризует газообмен, эффективность внешнего дыхания.
МВЛ=( ДО-АМП) х ЧД
3,5-4,5
л/мин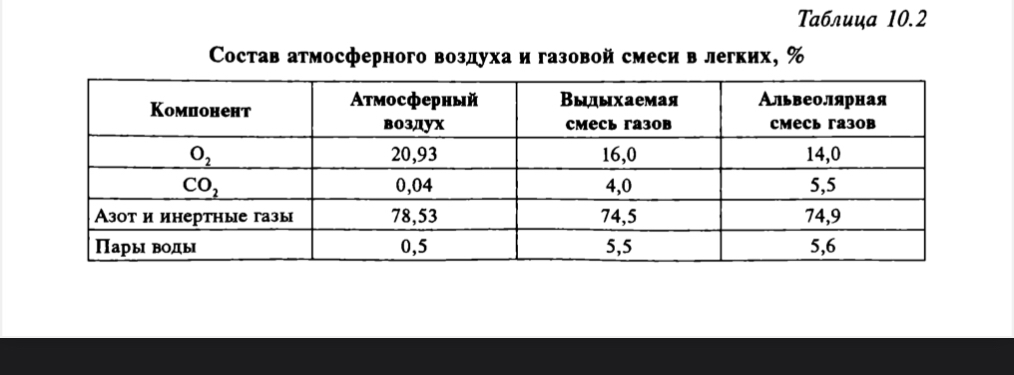
Легочная диффузия – пассивный процесс.
Факторы, способствующие диффузии в тканях лёгких:
Движущая сила, обеспечивающая давление, это градиент парциального давлени.
Большая скорость диффузии (выравнивания парциального давления кислорода в альвеолах и в крови лёгких происходит за 0,25с, кровь в капиллярах находится 0,5 секунд, то есть в 2 раба больше. Скорость диффузии углекислого газа в 20 раз больше таковой кислорода, так как он быстрее диффундирует, что обусловленно его лучшей растворимостью.)
Свойства газа. (Углекислый газ в 20-25 раз диффундирует быстрее, чем кислород, что обусловлено лучшей его растворимостью в жидкости и в мембранах.)
Большая диффузионная поверхность ( контакта легочных капилляров и альвеол 60-120м*2)
Корреляция между кровотоком в данном участке легкого (Q) и его вентиляцией (V).( Если участокплохо вентилируется, то кровеносные сосуды в этой области суживаются и закрываются, это осуществляется с помощью механизмов местной саморегуляции. Благодаря этому поддерживается отношение объема вентиляции лёгких к объёму кровотоков них в норме = 0,8-0,9.
Интенсивность вентиляции и кровообращения различных отделов легких, которая зависит от положения тела : в вертикальном положении лучше вентилируются нижние отделы, в горизонтальном – отделы легких, находящиеся внизу, в положение на спине – дорсальные. Примерно также изменяется кровообращение легких.
Скорость диффузии прямо пропорциальна площади поверхности барьера и градиенту парциального давления газа и обратно пропорциональна толщине барьера.

Кислородная ёмкость крови – это количество кислорода, которое может быть связано с гемоглобином у муж – 180-200мл/л; у жён – на 10-20% меньше.
Главный фактор связывания кислорода с гемоглобином – высокое парциальное давление кислорода в альвеолах – 100 мм рт.ст, в венозной крови парциальное давление = 40 мм рт ст, а в артериальной как в альвеолах.
Вспомогательные факторы: отщепление от карбогемоглобина углекислого газа и удаление его; понижение температуры в легких, увеличение рН крови.
Коэффициент утилизации кислорода – это процентное отношение доли кислорода, использованной тканями (разности концентраций кислорода в артериальной и венозной
Крови в % об.), к его концентрации в артериальной крови (в % об.).
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
.Каждый литр крови, содержащий 180-200 мл кислорода, отдаёт тканям 45мл кислорода – артериовенозная разницапо кислороду. Количество поглощенного кислорода – коэффициент использования кислорода.
78. Показатели внешнего дыхания: легочные объемы и емкости. Методы исследования внешнего дыхания у человека. Типы нарушения вентиляции легких: обструктивный и рестриктивный. Изменения показателей внешнего дыхания при нарушениях вентиляции легких.
Показатели внешнего дыхания:
Легочных объёмы:
Дыхательный объем (ДО) – количество воздуха, который человек вдыхает и выдыхает при спокойном дыхании(500 мл).
•Резервный объем вдоха (Рвд)– количество воздуха, которое человек может дополнительно вдохнуть после спокойного вдоха ( 1500-2500 мл).
•Резервный объем выдоха (Рвыд) – количество воздуха, которое человек может дополнительно выдохнуть после спокойного выдоха ( 1200-1500 мл).
Легочных ёмкости:
Жизненная емкость легких – наибольшее количество воздуха, которое человек может выдохнуть после максимального вдоха
3500-5000 мл
ЖЕЛ= ДО+РОвдоха+Ровыдоха
Функциональная остаточная емкость – количество воздуха, остающееся в легких после спокойного выдоха (2000-3000 мл.).
Это «буфер» между атмосферным и альвеолярным воздухом. Предотвращает выраженные колебания Р о2 и Р со2 в альвеолярном воздухе в течении дыхательного цикла
Емкость вдоха (Ев) – максимальное количество воздуха, которое можно вдохнуть после спокойного выдоха (2000-3000 мл).
Общая емкость легких (ОЕЛ) – количество воздуха, содержащееся в легких после максимального вдоха (4000-6000 мл.)
Остаточный объем (ОО)– количество воздуха, остающееся в легких после максимального выдоха (1000-1500 мл).
Методы исследования внешнего дыхания:
Спирометрия – регистрация легочных объемов (ДО, Ровд и выд, ЖЕЛ)
Спирография – графическая регистрация объемов воздуха, проходящих через легкие (ДО, МОД, МВЛ, ЖЕЛ, РО вд и выд, ИТ);
Пневмотахография – оценка объемной скорости дыхания.
Пнёвмография – графическая регистрация движения грудной клетки во время дыхания (не позволяет оценить легочные объемы)
Типы нарушений легочной вентиляции.
Рестриктивный тип - снижение дыхательных экскурсий легких (разница обхвата легких при вдохе и выдохе).
Причины:поражения легочной паренхимы (фиброз легких, пневмокониоз).
Признак: снижение ЖЕЛ, МВЛ
Обструктивный тип - сужение воздухоносных путей, повышение их аэродинамического сопротивления.
Причины:накопление в дыхательных путях слизи, набухание слизистой оболочки дыхательных путей, спазм бронхиальных мышц.
Признак: снижение индекса Тиффно, МВЛ
Показатели, их характеризующие.
Тест Тиффно- объём форсированного выдоха за секунду.
Индекс Тиффно = ОФВ1/ ЖЕЛ*100%.
В норме ИТ= 70-80%
Показатели: ЧД, МОД, МВЛ.
79. Регуляция внешнего дыхания: дыхательный центр, его структура. Роль периферических и центральных хеморецепторов, рецепторов растяжения легких, ирритантных, юкстакапиллярных рецепторов и мышечных веретен в регуляции параметров дыхания.
Дыхательный центр - совокупность взаимно связанных нейронов ЦНС, обеспечивающих координированную ритмическую деятельность дыхательных мышц и постоянное приспособление внешнего дыхания к изменяющимся условиям внутренней и внешней среды.
• Структуры, ответственные за процесс вдоха и выдоха, находятся в бульбопонтийной области мозга.
Дыхательный центр выполняет две основные функции в системе дыхания:
⎯ моторную, или двигательную, регулирующую сокращения дыхательных мышц
⎯ гомеостатическую, изменяющую характер дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Выделяют следующие части дыхательного центра.
1. Высшие отделы, расположенные выше ствола головного мозга, организуют взаимодействие нижележащих частей дыхательного центра, управляют произвольным дыханием и организуют взаимодействие системы дыхания с другими системами.
2. Пневмотаксический центр переднего моста (с апнейстическим центром) организует взаимодействие двух других структур следующего, нижнего уровня.
3. Медуллярные инспираторный и экспираторный центры (центры вдоха и выдоха), в соответствии с названием, посредством пейсмекернойритмической активности обеспечивают чередование вдоха и выдоха. Это происходит путем организации взаимодействия элементов нижележащего уровня (иерархии структур) дыхательного центра.
4. Мотонейроны спинного мозга, аксоны которых образуют нервы, идущие к дыхательным мышцам. Сами по себе мотонейроны не обладают способностью к осуществлению целостных актов дыхания. Дыхание возможно только при наличии связей совокупности мотонейронов спинного мозга, по крайней мере, с инспираторным и экспираторным центрами продолговатого мозга. При нарушении связей с отделами, расположенными выше продолговатого мозга, ухудшаются приспособительные возможности дыхания. Причем, чем выше локализация нарушения, тем более тонкие адаптационные возможности дыхания утрачиваются.
Центральные хеморецепторы расположены на вентральной поверхности продолговатого мозга и чувствительны к уровню углекислого газа и водородных ионов спинномозговой жидкости. Обеспечивают возбуждение дыхательных нейронов, т.к. поддерживают постоянный афферентный поток и участвуют в регуляции частоты и глубины дыхания при изменении газового состава спинномозговой жидкости.
Периферические рецепторы локализованы в области бифуркации сонной артерии и дуги аорты в специальных гломусах (клубочках). Афферентные волокна идут в составе блуждающего и языкоглоточного нервов в дыхательный центр. Реагируют на снижение напряжения кислорода, повышение уровня углекислого газа и водородных ионов в плазме крови.
Значение: обеспечивают рефлекторное усиление дыхания при изменении газового состава крови.
Вторичночувствующие рецепторы, сосудистые, неадаптирующиеся, всегда активны, увеличивается при изменениях.
Особенно сильным стимулом для хеморецепторов является сочетание гиперкапнии и гипоксемии. Это естественные сдвиги газового состава крови при физической нагрузке, которые приводят к рефлекторному увеличению легочной вентиляции.
Гиперкапния — повышение напряжения углекислого газа в плазме крови.
Гипоксемия — понижение напряжения кислорода в плазме крови.
При гипоксемии рост [H+] в ткани гломусов снижает проницаемость К-каналов мембраны рецепторов → деполяризация → открытие потенциалзависимых Са-каналов и диффузия ионов Сa внутрь клетки.
Са → экзоцитоз ДОФА. В области контакта мембраны рецептора с окончанием чувствительного нервного волокна → активность в волокнах синокаротидного нерва (нерв Геринга — часть языкоглоточного) → к ДЦ через нейроны ядер одиночного пути → рост вентиляции легких.
Рецепторы растяжения легких локализованы в гладкомышечном слое воздухоносных путей (трахея, бронхи), связаны толстыми афферентными миелиновыми волокнами с нейронами дыхательного центра, проходят в составе блуждающего нерва. При вдохе легкие растягиваются и активируются рецепторы растяжения легких, импульсы идут в дыхательный центр, вдох тормозится, а выдох стимулируется. Если перерезать блуждающие нервы, дыхание становится более редким и глубоким.
Значение: регулируют частоту и глубину дыхания, при спокойном дыхании не активны; низкопороговые.
Мощной рефлексогенной зоной является паренхима легких, обеспечивающая не только альвеолярное дыхание, но и рефлекторную регуляцию внешнего дыхания.
Основные типы легочных вагусных афферентов включают: медленноадаптирующиеся рецепторы растяжения альвеол, быстроадаптирующиеся рецепторы, С-волокна.
Важная роль в рефлекторной регуляции дыхания отводится проприорецепторам суставов грудной клетки, межреберных мышц, диафрагмы, сухожильным рецепторам. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которая через α-мотонейроны повышает активность α-мотонейронов и дозирует таким образом мышечное усилие.
В регуляции активности бульбарного дыхательного центра и внешнего дыхания принимает участие и афферентация с висцеральных рецепторов и рецепторов кожи, о чем свидетельствует развитие гипервентиляции легких при болевом и термическом раздражении.
B эпителии и субэпителиальном слое воздухоносных путей располагаются рецепторы, получившие название ирритантных. Особенно их много в области корней легких.
Для них характерными являются свойства как механорецепторов, так и хеморецепторов.
Возбуждаются они при очень сильных изменениях (увеличении или уменьшении) объема легких. Порог возбуждения них выше, чем всех остальных рецепторов. Импульсы в афферентных волокнах ирритантныхрецепторов возникают пачками только в течение короткого времени во время изменения объема легких, но часть из них возбуждается и при обычном вдохе и выдохе.
В качестве раздражителей ирритантных рецепторов могут быть:
· пылевые частицы;
· слизь;
· пары едких веществ (табачный дым, аммиак и др.);
· биологически активные вещества, образующиеся в стенках воздухоносных путей (гистамин).
· Они могут сильно раздражаться при ряде заболеваний (отек легких, пневмотораксе, бронхиальной астме и др.).
Раздражение ирритантных рецепторов приводит к:
· возникновению кашля;
· неприятным ощущениям типа жжения или першения;
· усилению инспираторной активности;
· укорачению фазы выдоха;
· увеличению частоты дыхания;
· рефлекторной бронхоконстрикции
J-рецепторы (юкстакапиллярные, юкстаальвеолярные) располагаются вблизи капилляров малого круга кровообращения в интерстициальной ткани альвеол. Афферентные волокна от них относятся к типу С. Точный механизм их участия в регуляции дыхания пока не ясен. Но, полагают, что основным раздражителем I-рецепторов является увеличение объема интерстициальной жидкости в легочной ткани.
Высокая активность J-рецепторов отмечается при:
· пневмониях;
· отеке легких;
· эмболии мелких сосудов легких;
· застое крови в малом круге кровообращения.
Раздражение J-рецепторов приводит к частому и поверхностному дыханию (тахипноэ - одышка) и бронхоконстрикции. В этих процессах наряду с J-рецепторами принимают участие и ирритантные рецепторы.
80. Транспорт кислорода кровью: роль эритроцитов и плазмы, кислородная емкость крови, коэффициент утилизации кислорода. Кривая диссоциации оксигемоглобина; факторы, влияющие на степень диссоциации оксигемоглобина.v
Транспорт О2 осуществляется в физически растворенном и химически связанном виде.
Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2.
Наиболее оптимальным является механизм транспорта О2 в химически связанном виде.
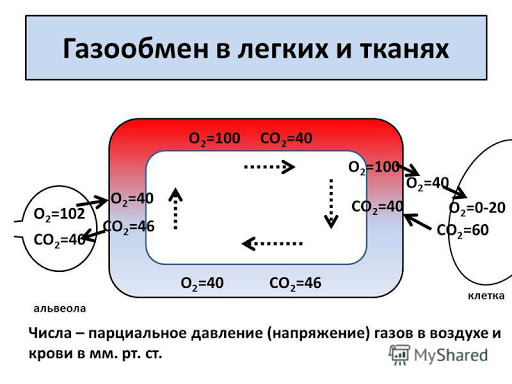
Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами.
В альвеолах легких парциальное давление О2 составляет 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2 составляет примерно 40 мм рт.ст.
Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2. Градиент О2 на альвеолярно-капиллярной мембране, равный в среднем 60 мм рт.ст., является одним из важнейших, но не единственным, согласно закону Фика, факторов начальной стадии диффузии этого газа из альвеол в кровь.
Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином.
Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются, и он может выполнять свою функцию на протяжении длительного времени.
Гемоглобин переносит О2 от легких к тканям.
Эта функция зависит от двух свойств гемоглобина:
1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 à НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном Рог в альвеолярном воздухе;
2) способности отдавать О2 в тканях (НbО2 à Нb + О2) в зависимости от метаболических потребностей клеток организма.
Кислородная емкость крови - это количество кислорода, связанного с гемоглобином, находящимся в 100 мл крови, при его полном насыщении кислородом.
Кислородную емкость крови определяют по концентрации Нb с двухвалентным железом (Fe2+). Максимально 1 моль гемоглобина может присоединить 4 моля О2 к своим 4 молям железа гема.
КУО2 -это процентное отношение доли кислорода, используемой тканями (разности концентраций кислорода в артериальной и венозной крови), к общему содержанию его в артериальной крови:
КУО2 = VO2а- VO2в / VO2а x 100 %
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 25-35%). Это позволяет тканям приспосабливаться к снижению доставки кислорода
Зависимость степени оксигенации гемоглобина от парциального давления Огв альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой. Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях.


На сродство кислорода к гемоглобину влияют различные метаболические факторы, что выражается в виде смещения кривой диссоциации влево или вправо.
Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата.
Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2) (см. рис. 8.7, А). Например, рН в эритроцитах на 0,2 единицы ниже, чем в плазме крови. В тканях вследствие повышенного содержания СО2 рН также меньше, чем в плазме крови. Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора».
Рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей или содержания 2,3-дифосфоглицерата вызывает сдвиг влево кривой диссоциации оксигемоглобина.
Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма.
Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2.