

31. Парасимпатический отдел автономной (вегетативной) нервной системы: особенности строения, медиаторы (нейротрансмиттеры), роль в регуляции функций организма.
Парасимпатическую иннервацию получают не все органы-ее лишены большинство кровеносных сосудов (кожи, мышц, мозга, органов брюшной полости), скелетные мышцы, мозговое вещество надпочечников.
Деятельность вегетативной нервной системы не зависит от воли человека. Однако все вегетативные функции подчиняются центральной нервной системе, в первую очередь — коре больших полушарий.
Функции:
нервная регуляция функций всех органов и тканей организма (кроме скелетных мышц);
регуляция обмена веществ;
поддержание гомеостаза организма;
приспособительные реакции всех позвоночных.
Особенности вегетативной нервной системы:
Очаговое расположение в мозге вегетативных нервных центров;
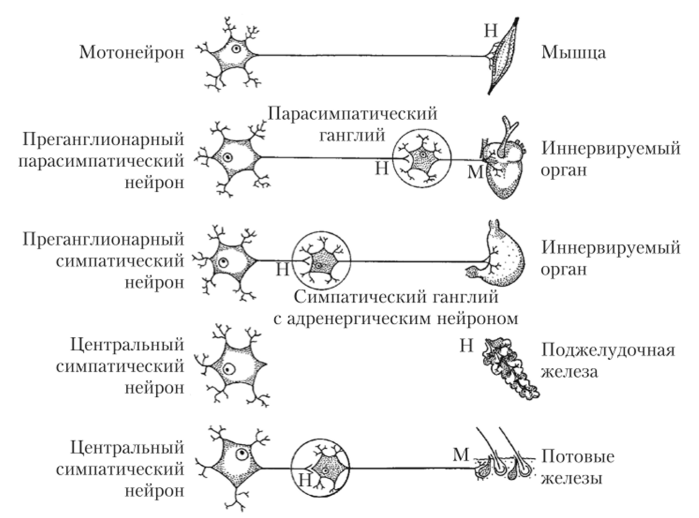
Эффекторные (двигательные) нейроны расположены за пределами центральной нервной системы в узлах вегетативных нервных сплетений;
Двухнейронный эфферентный нервный путь от мозга до рабочего органа;
Преобладают немиелинизированные нервные волокна, т.е. скорость проведения нервных импульсов ниже, чем в соматической нервной системе.
Парасимпатические ганглии расположены внутри органов, а в области головы-близко к органам. От их нервных клеток идут постганглионарные парасимпатические волокна, иннервирующие эффекторные клетки органов.
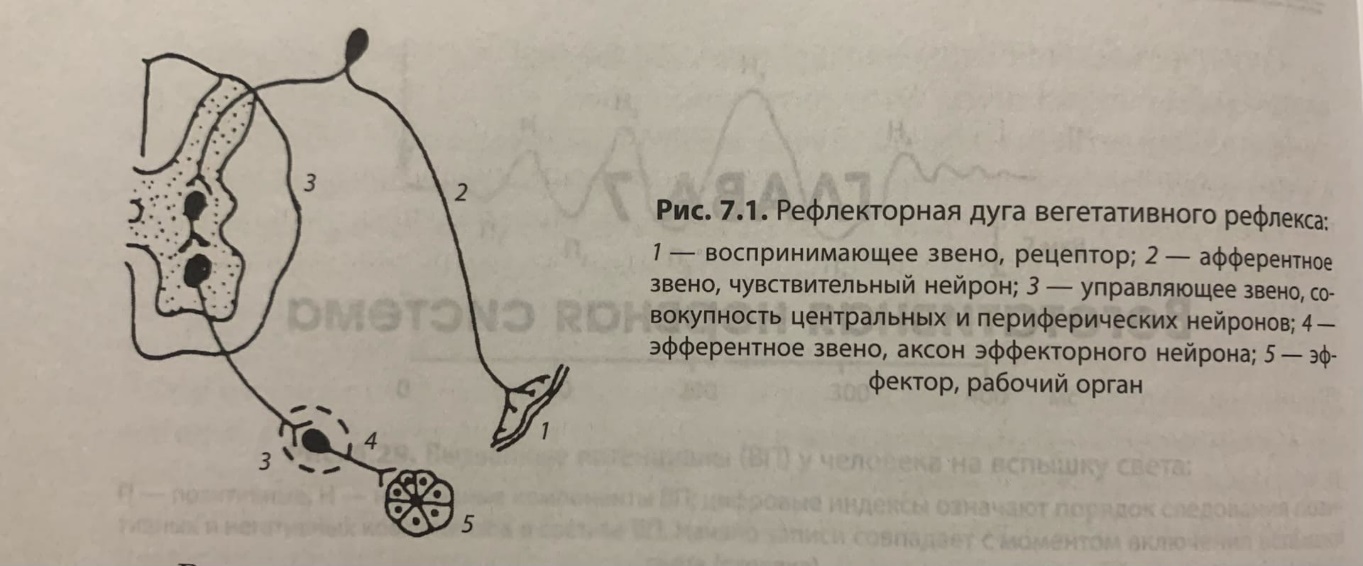
Центральную часть вегетативной нервной системы образуют вегетативные ядра — тела нейронов, лежащих в спинном и головном мозге. Они осуществляют координацию работы всех трех частей вегетативной нервной системы.
Периферическую часть вегетативной нервной системы образуют отходящие от ядер нервные волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов.
Парасимпатические нервные волокна имеются в черепных нервах (III, VII, IX, X пары) и в тазовом нерве (голубой цвет на схеме). Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамуса.
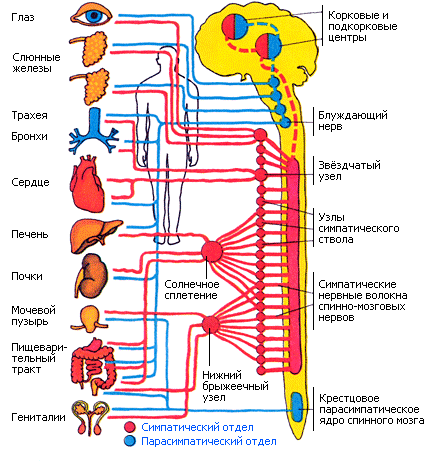
Парасимпатические волокна III пары ЧМН (глазодвигательный нерв) иннервируют сфинктер зрачка и цилиарную мышцу глаза, тем самым регулируя диаметр зрачка и степень аккомодации. Добавочное ядро (ядро Якубовича) локализовано в мезэнцефальном отделе среднего мозга.
Парасимпатические волокна VII пары ЧМН (лицевой нерв), его ветви, большой каменистый нерв-иннервирует слизистую носа, неба, слезную железу; барабанная струна-смешанный нерв, содержит чувствительные и секреторные волокна подчелюстной и подъязычной слюнных желез.
Парасимпатические волокна IX пары ЧМН (языкоглоточный нерв) подходят к околоушной железе в составе ушновисочного нерва-от 3 ветви тройничного нерва.
Преганглионарные волокна блуждающего нерва (X пары) начинаются от дорсального и двойного ядер продолговатого мозга. Дорсальное ядро иннервирует глотку, гортань, трахею, бронхи, сердце, пищевод, печень, поджелудочную и большую часть ЖКТ. Двойное ядро иннервирует сердце и поперечно-полосатые мышцы глотки, гортани, пищевода и мягкого неба.
Парасимпатические нервы сакрального отдела S2-S4 (тазовый нерв) иннервируют нисходящую часть ободочной кишки и тазовые органы-прямую кишку, мочевой пузырь и половые органы.
Медиаторы и рецепторы парасимпатического отдела
Медиатор преганглионарных и постганглионарных волокон-ацетилхолин (АХ). Он действует на инотропные N-холинорецепторы постсинаптической мембраны ганглионарного нейрона. Пузырьки с медиатором находятся в движении, его высвобождению способствуют ионы Са2+. АХ синтезируется в цитоплазме окончаний холинергических нейронов, депонируется в везикулах. Инактивация АХ происходит в синапсе с помощью ацетилхолинэстеразы с образованием холина и уксусной кислоты, идущей обратно на синтез АХ.
АХ реализует свое влияние посредством М-холинорецепторов:
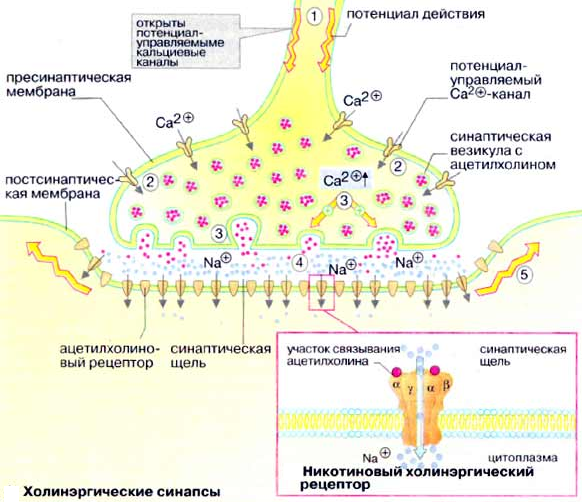
Они делятся на М1-, М2-, М3-холинорецепторы:
М1-холинорецепторы локализуются в вегетативных ганглиях.
М2-холинорецепторы-в сердце, на пресинаптических симпатических и парасимпатических окончаниях, в гладких мышцах ЖКТ.
М3-холинорецепторы-гладкие мышцы ЖКТ, большинство эндокринных желез.
Имеются М-холинорецепторы также в кровеносных сосудах, ЦНС.
К основным парасимпатическим влияниям относятся:
Органы |
Блуждающий нерв, рецепторы |
Все внутренние органы, кроме сердца |
Сокращение, М2-холинорецепторы |
Сфинктеры внутренних органов |
Расслабление, М3-холинорецепторы |
Сосуды |
НЕТ ИННЕРВАЦИИ |
Сердце |
Торможение, М2-холинорецепторы |
Секреция ЖКТ |
Стимуляция, М3-холинорецепторы |
Мужские половые гормоны |
Эрекция, М2-холинорецепторы |
Механизм действия ацетилхолина
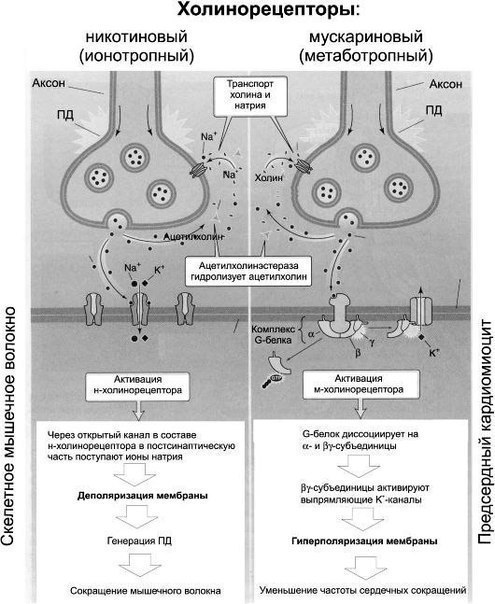
Большинство М-холинорецепторов активируются через посредников (например, в сердце, с помощью ИФ3) или через угнетение активности аденилатциклазы и синтеза цАМФ. При деполяризации клеток (=возбуждении) функция органа усиливается, при гиперполяризации-угнетается.
Возможна активация или инактивация К+ или Са2+-каналов с помощью ионотропных рецепторов. Пример: связывание АХ с М2-холинорецепторами пейсмекерных клеток сердца (они ионотропны) активирует К+-каналы через G-белок, гиперполяризацию этих клеток и как следствие замедление сердечной деятельности.
АХ, действуя на эндотелий сосуда, образует оксид азота NO-происходит расслабление гладких мышц сосуда, их расширение, что реализуется также через вторичных посредников-системы гуанилатциклаза-цГМФ, активирующей соответствующую протеинкиназу.
При избытке АХ развивается снижение возбудимости органа (десенситизация); при недостатке АХ-повышение возбудимости (сенситизация клеток-ффекторов).
32. Гуморальная регуляция (ауто-, пара- и эндокринная) функций внутренних органов. Гормоны, метаболиты, медиаторы, биологически активные вещества (определение, классификация). Механизмы их действия и роль в организме.
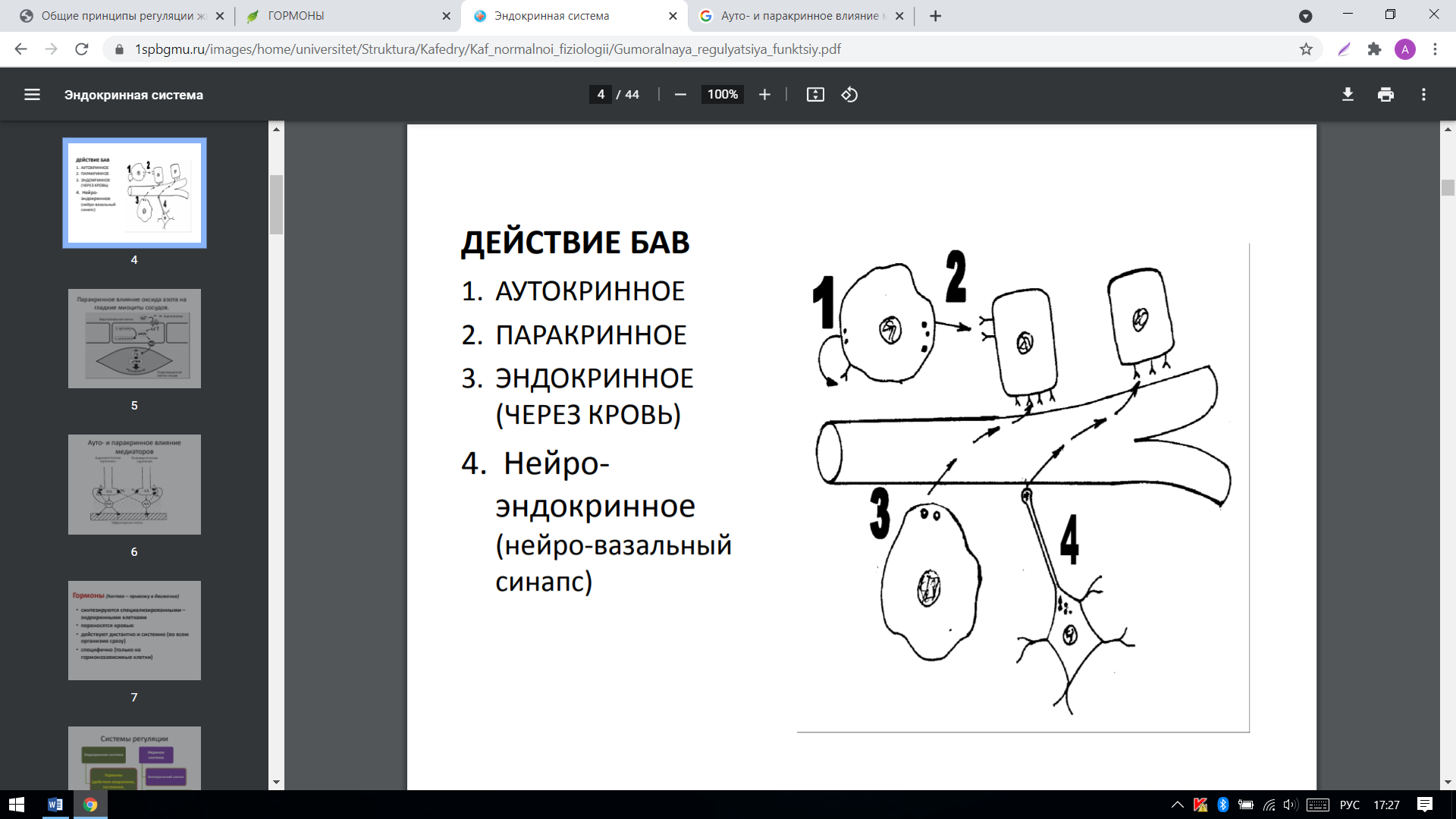
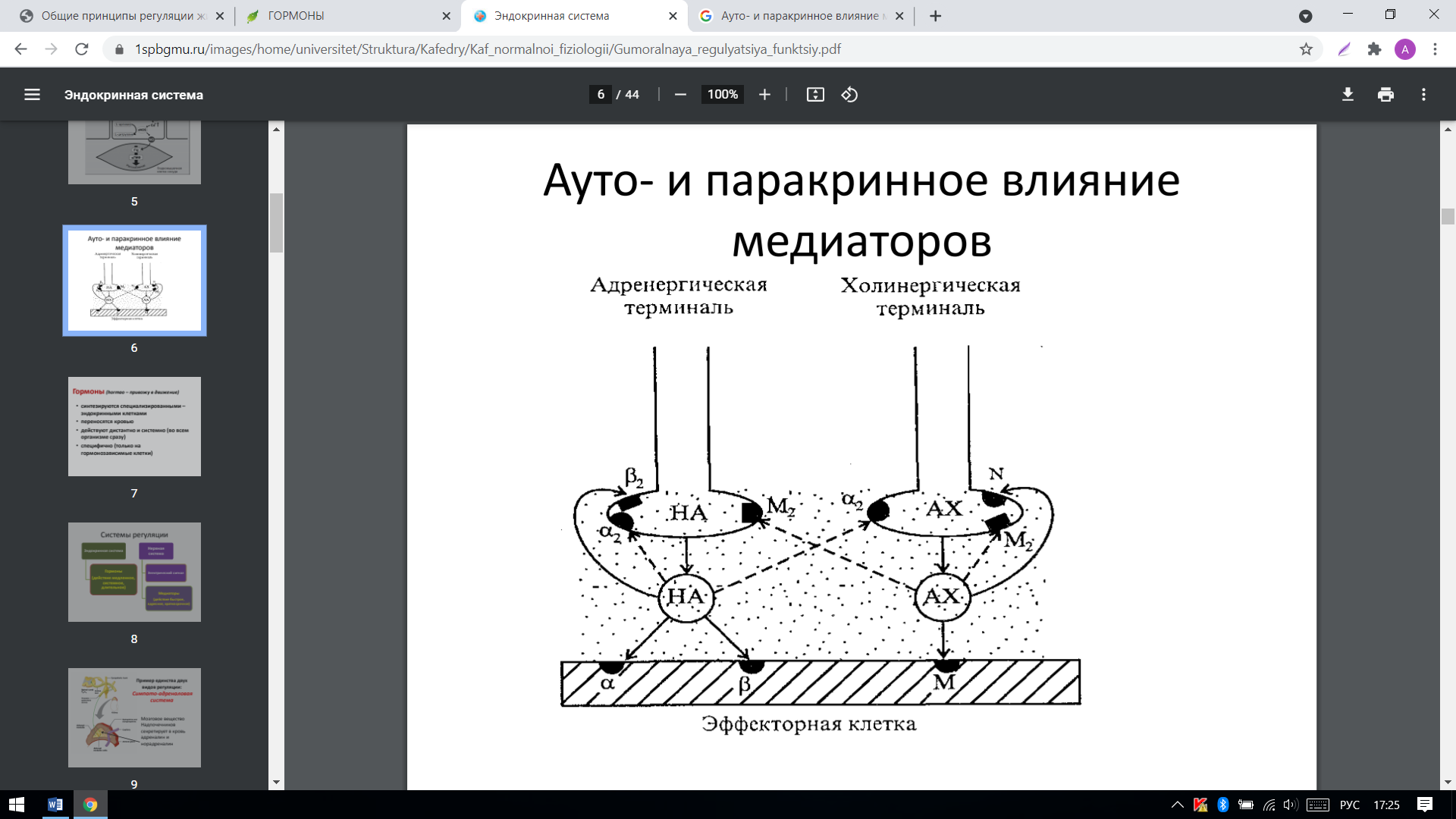
Аутокринное действие-действие БАВ преимущественно на рецепторы клетки, которая сама их и выделила.
Паракринное действие-действие БАВ на соседние клетки, минуя кровь.
Эндокринная система-это совокупность всех эндокринных клеток организма, которые способны продуцировать гуморальные факторы.
Звенья системы гормональной регуляции:
1. Звено управления (гипоталамус, гипофиз);
2. Звено синтеза и секреции гормонов;
3. Звено транспорта;
4. Звено депонирования;
5. Звено метаболизма;
6. Звено выделения;
7. Звено эффектора.
Гормоны – вещества, вырабатываемые специализированными клетками и регулирующие метаболические, морфогенетические и физиологические процессы. Тканевые гормоны (гуморальные факторы) вырабатываемые многими органами и тканями организма (гистамин, эйкозаноиды и др.).
Гормоны характеризуются: специфичностью действия; высокой биологической активностью; местным и дистантным действием; относительно небольшим периодом жизненного цикла (менее 1 ч).
Классификация гормонов рассмотрена подробно в следующем вопросе.
1. Мембранные рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве G-белка. G-белок влияет на ферменты, образующие вторичные мессенджеры (посредники). Последние передают сигнал на внутриклеточные белки. Большинство гормонов действуют по данному механизму:
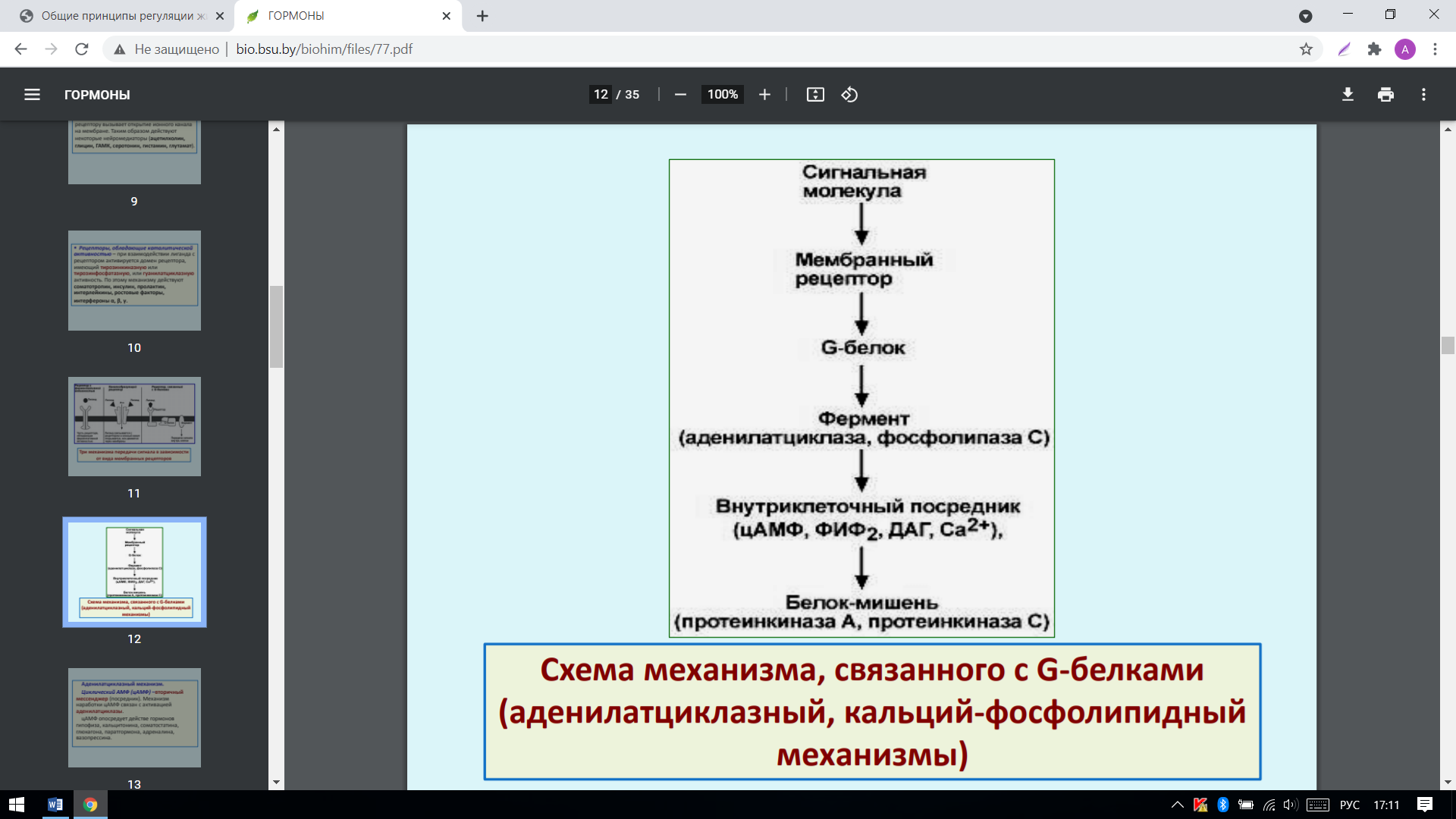
2. Выделяют также аденилатциклазный и кальций-фосфолипидный механизмы. Аденилатциклазный механизм: циклический АМФ (цАМФ) –вторичный мессенджер (посредник). Механизм наработки цАМФ связан с активацией аденилатциклазы. цАМФ опосредует действе гормонов гипофиза, кальцитонина, соматостатина, глюкагона, паратгормона, адреналина, вазопрессина:
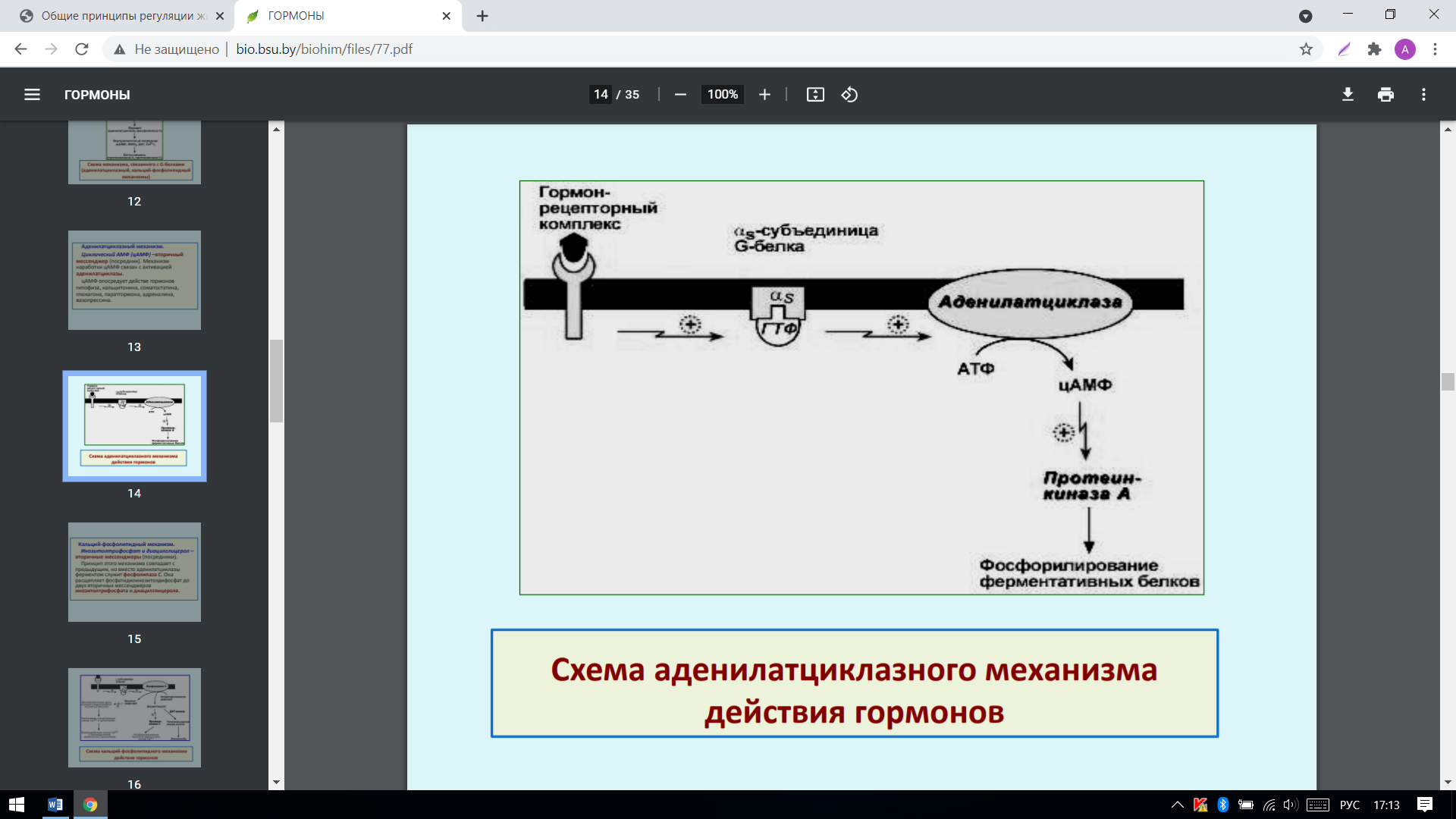
*Гуанилатциклазный механизм передачи сигнала схож с аденилатциклазным: лиганд взаимодействует непосредственно с ферментом – гуанилатциклазой, которая нарабатывает цГМФ (вторичный мессенджер). цГМФ воздействует на протеинкиназу G, которая фосфорилирует белки, влияющие на метаболизм клетки. По этому механизму действуют соматотропин, инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ.
3. Кальций-фосфолипидный механизм: Инозитолтрифосфат и диацилглицерол – вторичные мессенджеры (посредники). Принцип этого механизма совпадает с предыдущим, но вместо аденилатциклазы ферментом служит фосфолипаза С. Она расщепляет фосфатидилинозитолдифосфат до двух вторичных мессенджеров инозитолтрифосфата и диацилглицерола:
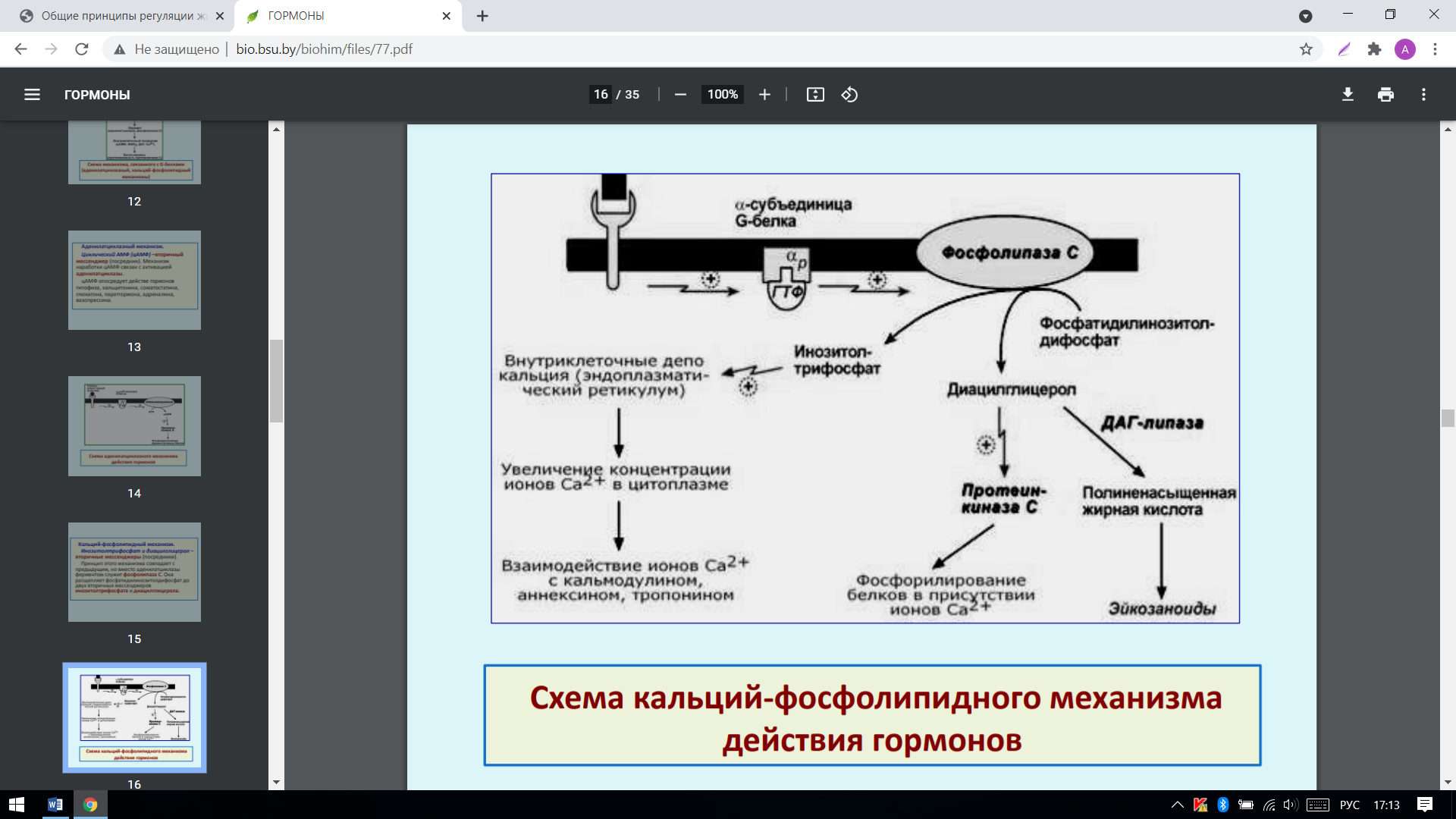
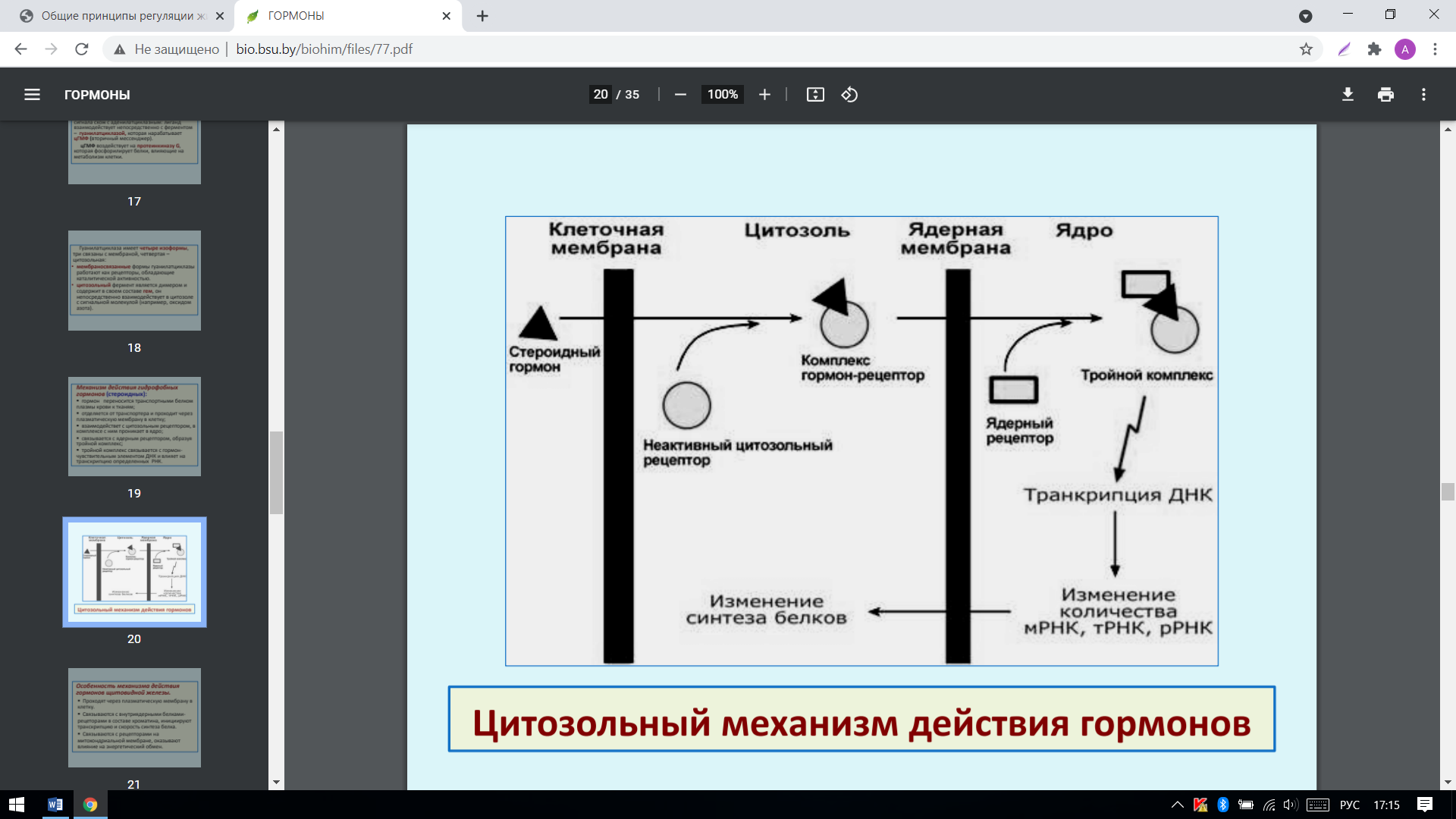
Механизм действия гидрофобных гормонов (стероидных):
гормон переносится транспортными белком плазмы крови к тканям;
отделяется от транспортера и проходит через плазматическую мембрану в клетку;
взаимодействет с цитозольным рецептором, в комплексе с ним проникает в ядро;
связывается с ядерным рецептором, образуя тройной комплекс;
тройной комплекс связывается с гормончувствительным элементом ДНК и влияет на транскрипцию определенных РНК:
Нейромедиаторы
Нейромедиаторы (нейротрансмиттеры, медиаторы) — биологически активные химические вещества, посредством которых осуществляется передача электрохимического импульса от нервной клетки через синаптическое пространство между нейронами, а также, например, от нейронов к мышечной ткани или железистым клеткам. Рецепторы, связанные с ионными каналами – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом действуют некоторые нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин, гистамин, глутамат).
Метаболиты
Метаболиты: ионы, глюкоза, кислород, углекислый газ (концентрация в плазме около 10-3 моль в литре). Поток метаболитов в обмене веществ определяется прежде всего активностью ферментов, особенно ключевых ферментов. Контроль за биосинтезом фермента осуществляется на генетическом уровне-синтез соответствующей мРНК, транскрипции кодирующего фермент гена.
Взаимопревращение ключевых ферментов: При метаболической потребности активирующий фермент (E1) переводит ключевой фермент в активную форму. Если потребность в этом пути обмена веществ отпадает, инактивирующий фермент (E2) снова переводит ключевой фермент в неактивную форму.
Важными параметрами, контролирующими протекание метаболического пути, являются потребность в первом метаболите и в доступности кофермента. Доступность метаболита возрастает с повышением активности метаболического пути, в котором он образуется, и падает с повышением активности других путей, в которых он расходуется.
Ингибируется ключевой фермент конечными продуктами реакции (по типу обратной связи), активировать фермент может первый метаболит цепи реакций.