

7. Потенциал действия и его фазы. Изменение возбудимости в процессе возбуждения. Рефрактерность, ее виды и причины.
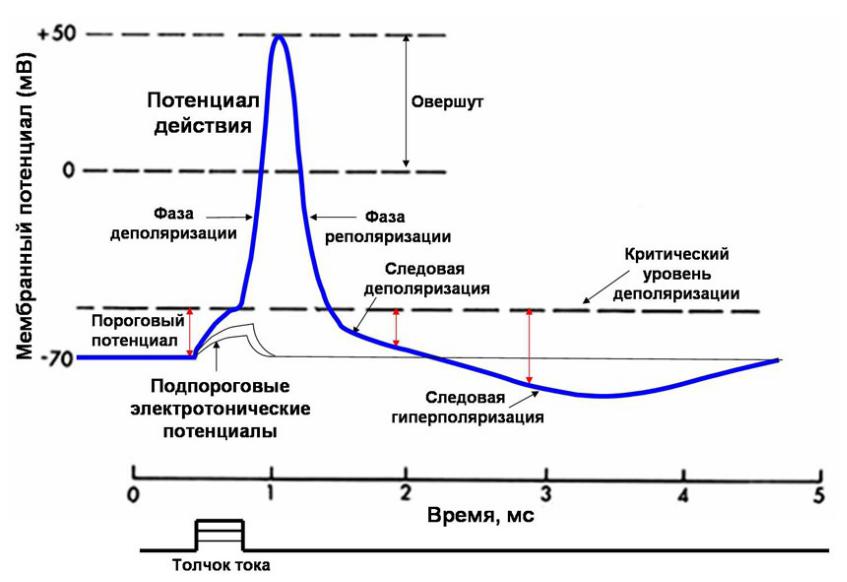
Потенциал действия – быстрое изменение мембранного потенциала, которое быстро распространяется вдоль клетки.
Каждый ПД начинается со сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу.
Фазы ПД:
Стадия покоя представлена мембранным потенциалом покоя, который предшевствует потенциалу действия. Поляризация мембраны -90мВ Действие раздражителя, возникновение локальных ответов, открытие медленных натриевых каналов, медленная деполяризация, достижение КУД
Быстрая деполяризация – лавинообразный вход Na в клетку, инверсия заряда мембраны (внутри +, а снаружи -), заряд мембраны уменьшается вплоть до противоположного (это называется «овершут»), закрываются натриевые каналы.
Реполяризация – в результате выхода калия из клетки, калий выходит с избытком и возникает следовая деполяризация, следовая гиперполяризация.
Восстановление покоя.
Изменение возбудимости в процессе возбуждения. Возбудимость клетки во время ее возбуждения быстро и сильно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и так же, как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов.
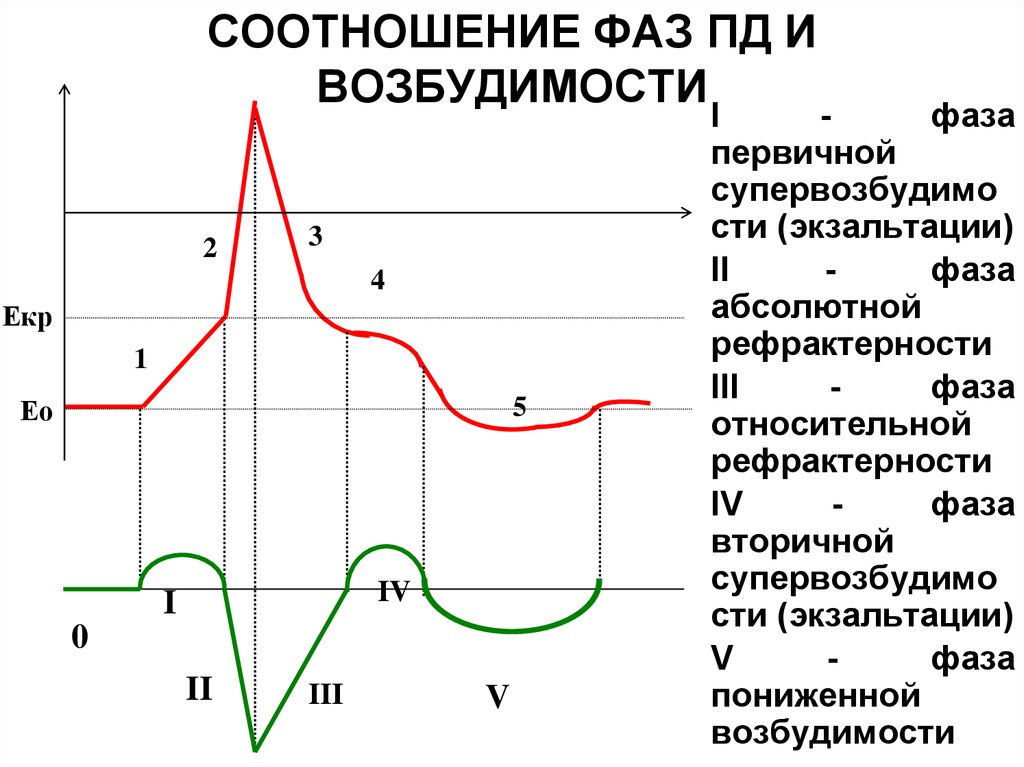
1. Кратковременное повышение возбудимости (первичной супервозбудимости) в начале развития ПД, когда уже возникла некоторая деполяризация клеточной мембраны (какое-то количество Na вошло в клетку). Возбудимость повышена, так как клетка частично деполяризована. Если в этот период добавить раздражителя, то возникнет ПД. (соответствует медленной деполяризации)
2. Абсолютная рефрактерная фаза— это полная невозбудимость клетки (возбудимость равна нулю), она соответствует пику ПД и продолжается 1—2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефрактерная фаза. Клетка в этот период времени на раздражения любой силы не отвечает. Это объясняется тем, что потенциал-зависимые натриевые каналы итак уже все открыты, натрий лавинообразно входит в клетку, вызывая деполяризацию, дополнительное раздражение клетки ничего не изменит. (соответствует быстрой деполяризации и чуть-чуть захватывает от реполяризации, на картинке этого не видно)
3. Относительная рефрактерная фаза— это период восстановления возбудимости клетки, когда сильное раздражение может вызвать новое возбуждение. Относительная рефрактерная фаза соответствует реполяризации. Пониженная возбудимость является следствием все еще повышенной проницаемости для К+ и избыточного выхода его из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, так как выход К+ из клетки препятствует деполяризации мембраны. (соответствует фазе реполяризации)
4. Фаза экзальтации (вторичной супервозбудимости)— это период повышенной возбудимости. Он соответствует следовой деполяризации. Очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Na+, Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность. (соответствует фазе следовой деполяризации)
5. Фаза пониженной возбудимости - в период следовой гиперполяризации мембранный потенциал больше (то есть ниже на графике) и дальше отстоит от критического уровня деполяризации., а значит, чтобы вызвать ПД раздражение должно быть сильнее. (соответствует фазе следовой гиперполяризации).
Итого, что мы имеем: мембрана клетки не возбудима в фазу абсолютной рефрактерности, мембрана более возбудима в фазу первичной и вторичной возбудимости, в фазу относительной рефрактерности возбудимость низкая, а в фазу пониженной возбудимости нужен сильный раздражитель, чтобы вызвать дьявола (ПД конечно-же).
8. Ионный механизм генерации потенциала действия. Роль ионных концентрационных градиентов в формировании потенциала действия. Состояние ионных каналов в различные фазы потенциала действия. Регистрация биопотенциалов (ЭЭГ, ЭКГ, ЭМГ и др.)
Ионы двигаются через каналы. На мембране имеются следующие каналы: K-Na насос, свободно проницаемые калиевые каналы (через них калий двигается из клетки по гардиенту концентрации, эти каналы всегда открыты), вольтаж-зависимые натриевые и калиевые каналы (реагируют на изменение мембранного потенциала и открываются, у этих каналов есть два вида ворот – снаружи активирующие ворота, изнутри - инактивирующие), лиганд-зависимые ионные каналы (открываться эти каналы будут при связывании с каким-либо лигандом (нейромедиаторы)).
Нейромедиатор связывается с рецептором и лиганд-зависимый канал становится открытым и проводимым для Na, тогда натрий по гардиенту концентрации будет заходить в клетку. Небольшое количество положительно-заряженных ионов теперь находится в клетке – это значит, что внутренний заряд в клетке становится менее отрицательным. Это соответствует медленной деполяризации. Допустим, -70 мВ – потенциал покоя клетки, теперь он стал -60мВ. Помним про вольтаж-зависимые каналы? Настало их время, ибо изменилась поляризация.
Вольтаж-зависимые каналы открылись (активирующие ворота открылись) и стали обладать проводимостью для Na – лавинообразный ток натрия по гардиенту концентрации, что соответствует быстрой деполяризации. Инактивирующие вороты пока что открыты и у натрия есть время побольше втиснуться в клетку. Инактивирующие ворота закрываются, активирующие открыты, но натрий не входит, а все уже, а надо было раньше. (вольтаж-зависимые натриевые каналы не могут из инактивированного состояния вернуться в активированное, для этого нужно, чтобы сначала закрылись активирующие ворота, потом открылись инактивирующие ворота – то есть канал должен вернуться в состояние покоя, а для этого нужно, чтобы внутри клетки был потенциал покоя (-90 в нашем примере) – этой особенностью объясняется период рефрактерности клетки, мы не сможем открыть канал заново, пока клетка не вернется к покою). Заряд в клетке положительный (овершут).
После закрытия вольтаж-зависимые натриевые каналы открываются калиевые. Далее калий начинает выходить из клетки через вольтаж-зависимые калиевые каналы, которые чувствительны к положительному заряду мембраны, открылись на пике деполяризации. К двигается по гардиенту концентрации из клетки через свои свободные каналы и через открытые для него вольтаж-зависимые. Это приводит к уменьшению положительно-заряженных частиц в клетке, движению к отрицательному заряду – реполяризация. При избыточном выходе калия возникает гиперполяризация.
Виды регистрации
1- Внутриклеточная монополярная (микроэлектроды) 2- Внеклеточная биполярная(ЭМГ,ЭКГ,ЭЭГ)
Электромиография (ЭМГ) потенциалов, возникающих в скелетных мышцах человека и животных при возбуждении мышечных волокон; регистрация электрической активности мышц.
Электроэнцефалография (ЭЭГ) — регистрация суммарной электрической активности мозга, отводимой с поверхности кожи головы, а также метод записи таких потенциалов.
Электрокардиография — методика регистрации и исследования электрических полей, образующихся при работе сердца.
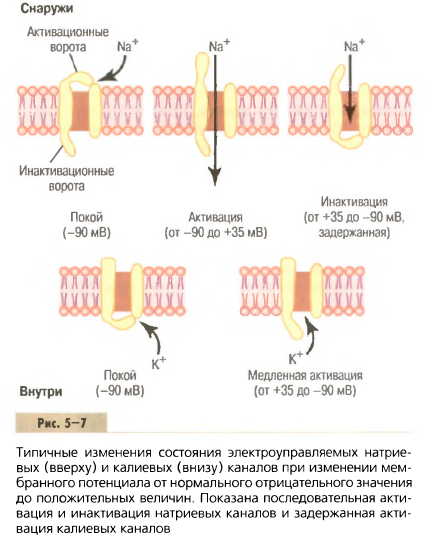
Вольтаж-зависимые=электроуправляемые
9. Физиологические свойства скелетных мышц. Нейромоторная (двигательная) единица. Виды двигательных единиц. Типы мышечных сокращений. Одиночное сокращение, его фазы. Суммация одиночных сокращений и тетанус. Сила и работа мышц.
Скелетная мускулатура является составной частью опорно-двигательного аппарата человека.
Физиологические свойства скелетной мышцы:
1) Возбудимость — способность отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала. В естественных условиях этим раздражителем является медиатор ацетилхолин, который выделяется в пресинаптических окончаниях аксонов мотонейронов. МП клеток скелетных мышц -90 мВ. Амплитуда ПД составляет 120-130 мВ. Скелетные мышцы обладают высокой чувствительностью к действию электрического тока.
2) Низкая проводимость (10-13 м/с) — способность проводить потенциал действия вдоль и вглубь мышечного волокна по Т-системе;
3) Сократимость — способность укорачиваться или развивать напряжение при возбуждении;
4) Эластичность — способность развивать напряжение при растягивании.
5) Рефрактерность – отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Занимает по времени больший отрезок, чем у нервного волокна.
6) Лабильность – функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях
Нейромоторная (двигательная) единица. Виды двигательных единиц.
Каждый мотонейрон, аксон которого покидает спинной мозг, иннервирует много мышечных волокон, количество которых зависит от типа мышцы. Все мышечные волокна, иннервируемые одиночным нервным волокном, называют моторной единицей.
Классификация двигательных единиц: (ДЕ I, ДЕ II-В, ДЕ II-A)
Медленные, неутомляемые двигательные единицы (ДЕ I) состоят и мотонейронов малого размера, имеющих низкий порог возбудимости, высокое входное сопротивление. При деполяризации мелких нейронов возникает продолжительный разряд с незначительной адаптацией. Мотонейроны с такими свойствами называются тоническими. Небольшой диаметр аксона (до 5 -7 мкм) объясняет и невысокую, по сравнению с более толстыми, скорость проведения возбуждения. Мышечные волокна, входящие в ДЕ этого типа, относятся к красным волокнам (тип I), имеющим наименьший диаметр, скорость их сокращения минимальна, максимальное напряжение слабее, чем белых волокон (тип II), они характеризуются малой утомляемостью.
Быстрые, легко утомляемые двигательные единицы (тип ДЕ II В) сформированы из крупных мотонейронов, имеющих высокий порог возбуждения, диаметр их аксонов наибольший (до 15 мкм), скорость проведения возбуждения достигает 120 м/с, высокочастотная импульсация кратковременна и быстро спадает, т.к. происходит быстрая адаптация. Крупные мотонейроны относятся к нейронам фазического типа. Входящие в эти ДЕ мышечные волокна относятся ко II типу (белые волокна). Они способны развивать значительное напряжение, но быстро утомляются.
Третий тип двигательных единиц - тип ДЕ II-A относится к промежуточному типу. В их состав входят как быстрые, так и медленные мышечные волокна. Мотонейроны - среднего калибра.
Скелетные мышцы, в зависимости от их функциональных особенностей, состоят из различного набора двигательных единиц. Тип ДЕ формируется в процессе онтогенеза и в зрелой мышце соотношение быстрых и медленных ДЕ уже не меняется. В целой мышце мышечные волокна одной ДЕ перемежаются с волокнами нескольких других ДЕ.
Типы мышечных сокращений. Одиночное сокращение, его фазы. Суммация одиночных сокращений и тетанус.
Мышечные сокращения бывают:
Фазные – кратковременные, быстрые, высокоамплитудные
Тонические – длительные, медленные, низкоамплитудные
Еще одна классификация мышечных сокращений:
Одиночное – при однократном раздражении нерва/мышцы
Тетанические – длительное сокращение при ритмичном действии раздражителя
Одиночное мышечное сокращение состоит из 3 фаз. При раздражении мышцы одиночным импульсом возникает одиночное мышечное сокращение.
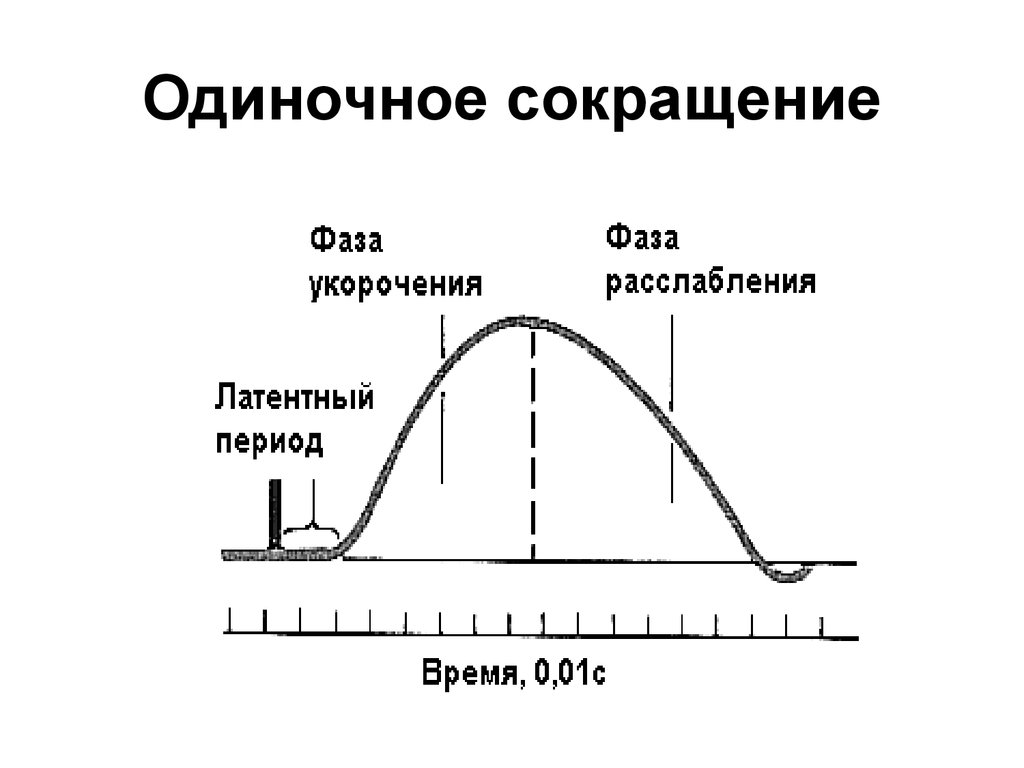
Латентный период – промежуток между моментами подачи стимула и началом сокращения (2мс)
Период укорочения или развития напряжения за счет скольжения актиновых и миозиновых нитей
Период расслабления, возникающий за счет уменьшения внутриклеточной концентрации ионов Са, отсоединения головок миозина от актиновых филаментов и эластичности.
Тетаническое сокращение (тетанус) – это длительное сокращение мышцы, возникающее под действием ритмического раздражения. Различают 2 вида тетануса: гладкий и зубчатый.
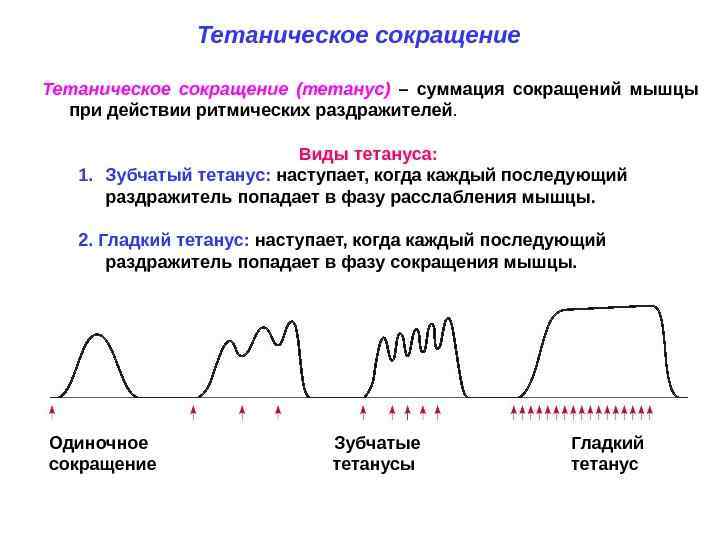
Обратите внимание, на картинке выше написано, а какой момент нужно действовать раздражителем, что бы получился тот или иной тетанус.
Сила и работа мышц.
Сила мышцы – максимальная масса груза, который может быть поднят при сокращении мышцы, выражается в ньютонах.
Сила мышцы зависит от: анатомического строения (суммы поперечных сечений всех волокон, из которых состоит мышца), исходной длины мышцы, числа возбужденных волокон (сила одиночного сокращения) и от тренированности мышцы.
Работа мышц – сила, затраченная мышцей на перемещение тела на определенное расстояние. Работа рассчитывается по формуле: A=F*L (Дж). Где F – сила приложенная к телу, L – расстояние перемещения тела.
Существует динамическая (сокращение мышц при движении) и статическая (сокращение мышц при поддерживании позы) работа.
В результате продолжительной деятельности работоспособность мышцы снижается – утомляемость. Сила сокращений при этом изменяется, длительность одного мышечного сокращения увеличивается. Наиболее выражено изменение длительности одиночного сокращения.
Динамометрия – измерение силы разных групп мышц
10. Механизм мышечного сокращения и расслабления. Электромеханическое сопряжение. Роль Са2+ в мышечном сокращении. Регуляторные и сократительные белки скелетных мышц. Гипертрофия и атрофия мышц. Проблема гиподинамии.
Смотрим на картиночку ниже)))
Сокращение: Генерация ПД на мембране мышечной клетки(1)→возбуждение мембраны Т- трубочек(2) → открытие Са++ каналов саркоплазматического ретикулума (СПР)(3) →выход Са++ в цитоплазму (4) → образование комплекса Са++ + тропонин (5) →смещение тропомиозина с активных центров актина → образование актомиозиновых мостиков → скольжение актина относительно миозина → укорочение мышцы .
Расслабление: Активация Са++ насоса СПР (6) → секвестрация Са++ в СПР → отсоединение Са++ от тропонина → возвращение тропомиозина на активные центры актина → блокирование образования актомиозиновых мостиков → восстановление исходной длины мышцы.
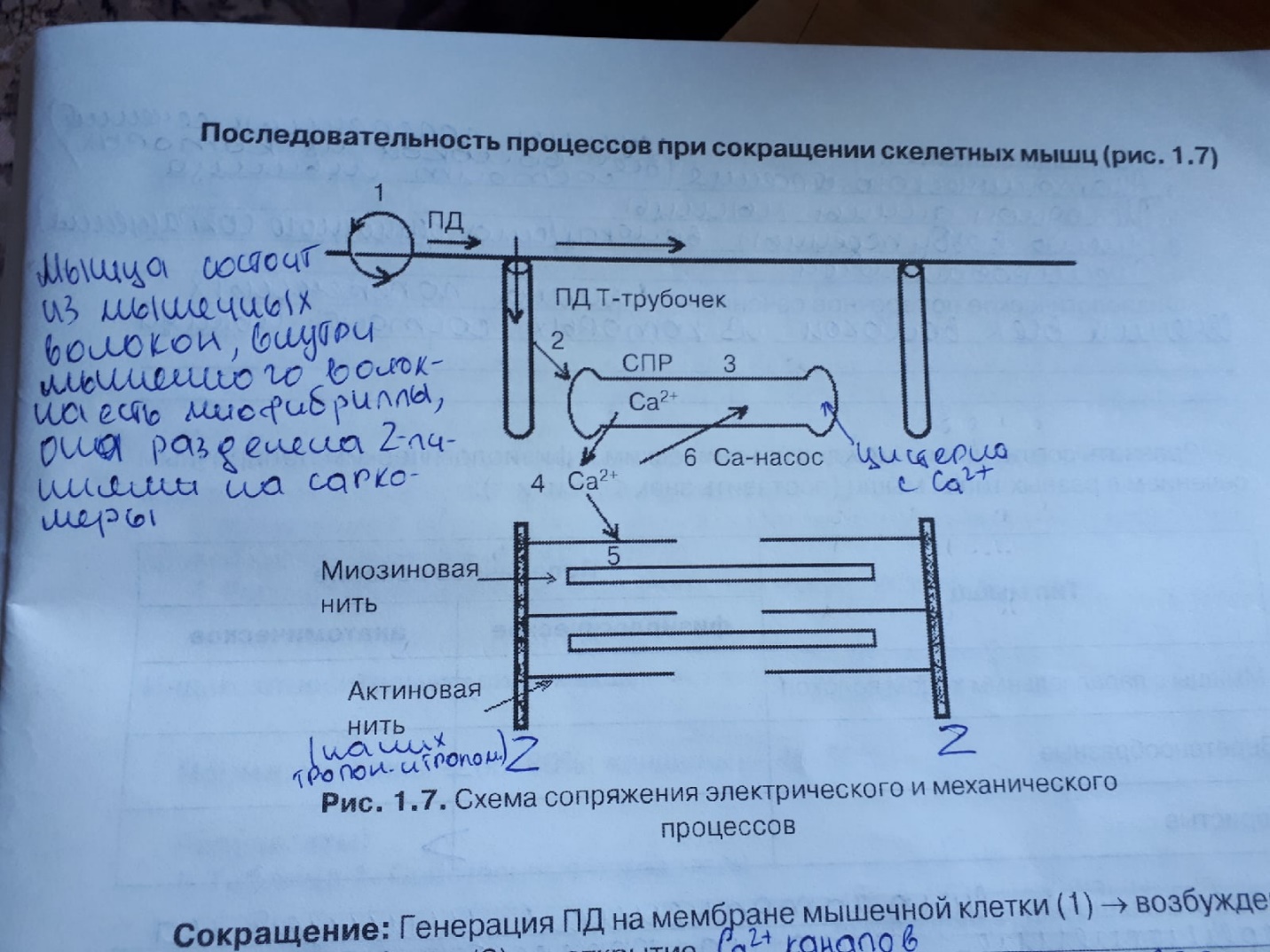
Электромеханическое сопряжение - это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску цикла поперечных мостиков.
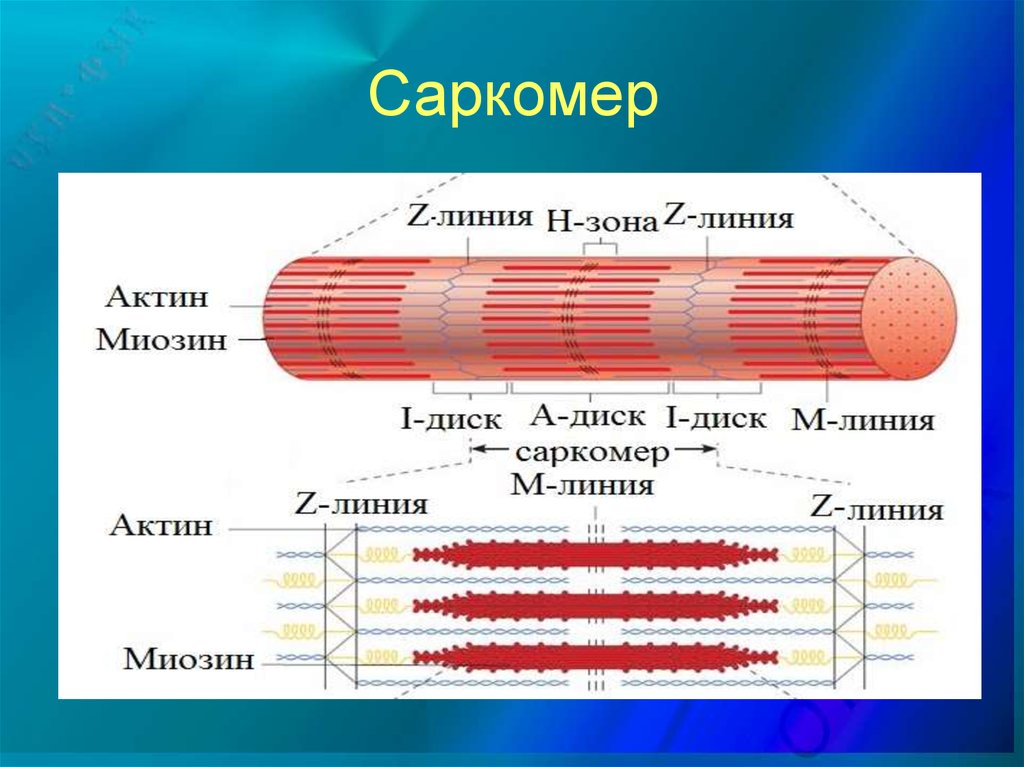
Мышца состоит из волокон. Волокно состоит из миофибрилл. Миофибрилла включает в себя последовательно соединенные белки – саркомеры. Саркомер является сократительной единицей мышечного волокна. Смотрим на картинку ниже и видим: А-диск – образован миозиновыми протофибриллами. Светлый участок саркомера между миозиновыми А-исками называется I-диском – он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок. Каждый саркомер имеет 2 набора тонких нитей, прикрепленных к Z- полоскам и один комплект толстых нитей, сосредоточенных в А-диске. Чередование участков актина и миозина придают мышце поперечную исчерченность.
Протофибриллы – актин и миозин – нити белка. Актиновые нити состоят из двух скрученных в спираль субъединиц, в состав актиновых нитей входят регуляторные белки – тропомиозин и тропонин.
Укорочение мышцы – результат сокращения множества саркомеров. При сокращении актиновые нити скользят относительно миозиновых, но длина нитей не изменяется.

Электромеханическое сопряжение.
Генерируется ПД в мышечном синапсе, возбуждение распространяется по мембране миоцита. Распространяется и на Т-трубочки (это такие штуки (инвагинации) от сарколеммы (оболочки, покрывающей мышцу) идут вглубь и соединяются с расположенными между миофибриллами цистернами, содержат кальций).
ПД Т-трубочек активирует открытие кальциевых каналов на саркоплазматическом ретикулюме (это те цистерны), кальций выходит из цистерн по гардиенту.
Са контактирует с тропонином, из-за этого смещается тропомиозин – освободилось местечко на актиновой нити – местечко для головки миозинового мостика.
Миозин соединяется с актином, образуя поперечный мостик. Расходуется АТФ на движение этого мостика, он двигается как весло в лодке, двигая лодку относительно морской глади – миозин относительно актина. Молекула АТФ присоединяется к головке миозина, отрывая миозин от актина. (то есть одна атф нужна для того, чтобы создалось движение и другая атф для отрывания головки)
Головки поперечных мостиков вовращаются в исходное положение и устанавливают контакт со следующим участком актина. – такие гребковые двиения повторяются многократно.
Расслабление мышцы происходит благодаря процессам, идущим в обратном направлении.
Последовательность событий, начиная от связывания поперечного мостика с тонким филаментом и до момента, когда система готова к повторению процесса, называется рабочим циклом поперечных мостиков. Каждый цикл состоит из четырех стадий:
- прикрепление поперечного мостика к тонкому филаменту;
- движение поперечного мостика, создающее напряжение тонкого филамента;
- отсоединение поперечного мостика от тонкого филамента;
Кальций способствует выбросу нейромедиатора в синаптическую щель (в нервно-мышечном синапсе) – тогда сгенерируется ПД на мембране миоцитов. Кальций освобождает активный центр актина. Кальций способствует расслаблению мышцы (кальциевая АТФ-аза)
Основную сократительную функцию во всех видах мышц осуществляют тонкие и толстые нити-миофиламенты (миофибриллы) актин и миозин.
Вспомогательную - регуляторную осуществляют тропомиозин (ТгМ, ММ:68 кО) и комплекс тропонина (Тг, ММ:70 кО), который состоит из субъединиц.
Увеличение общей массы мышцы называют мышечной гипертрофией, а уменьшение — мышечной атрофией.
Мышечная гипертрофия практически всегда является результатом увеличения количества актиновых и миозиновых нитей в каждом мышечном волокне, что ведет к их укрупнению. Это называют простой гипертрофией волокон. Степень гипертрофии значительно возрастает, если во время сокращения мышца нагружена.
Гиподинами́я — нарушение функций организма (опорно-двигательного аппарата, кровообращения, дыхания, пищеварения) при ограничении двигательной активности, снижении силы сокращения мышц. Распространённость гиподинамии возрастает в связи с урбанизацией, автоматизацией и механизацией труда, увеличением роли средств коммуникации.
Вопросы к экзамену по физе №11-15 (тут про гладкие мышцы, нервы, синапсы и рецепторы) P.s листай до конца.