

5. Сердце: источники развития, функции, строение стенки. Виды кардиомиоцитов, их функциональное значение и регенерация. Проводящая система сердца: строение и функциональное значение.
Развитие, ф-ии, стенку, сократительные кардиомиоциты, регенерацию смотреть выше.
Секреторные кардиомиоциты - лежат преимущественно в предсердии и ушках сердца, содержат гранулы с предсердным натрийуретическим фактором (ПНУФ)(пептидный гормон)
Давление повышается, в предсердие поступает много крови, предсердие растягивается, ПНУФ выходит в кровь, поступает к собирательным трубочкам почки и клубочковой зоне коры надпочечников, из организма выделяется вода, давление падает
Кроме того гипотензивный эффект
Проводящие - цитоплазма светлая, ядра крупные, сократительный аппарат плохо развит, волокна не упорядочены, поперечной исчерченности нет, Т-трубочек нет, Могут проводить электрический импульс
Выделяют - клетки-пейсмейкеры (Р-клетки, водители ритма) - лежат в основном в синоатриальном узле, способны к спонтанной деполяризации мембраны и генерации ПД. Он распространяется по проводящим кардиомиоцитам к рабочим через нексусы
Переходные (промежуточные) проводящие кардиомиоциты - нечто среднее между собственно проводящими и пейсмейкерами (могут и проводить и генерировать импульс, но генерируют его с меньшей частотой)(в атрио-вентрикулярном узле)
Собственно проводящие (клетки-волокна) - формируют пучок Гиса и волокна Пуркинье
В пучке Гиса- миофибриллы длинные, идут спирально, гликогена мало
В волокнах Пуркинье - миофибриллы неупорядоченные, гликогена много, самые крупные клетки миокарда, много щелевых контактов, соединены нексусами и десмосомами
Функция - проведение ПД
Проводящая система сердца
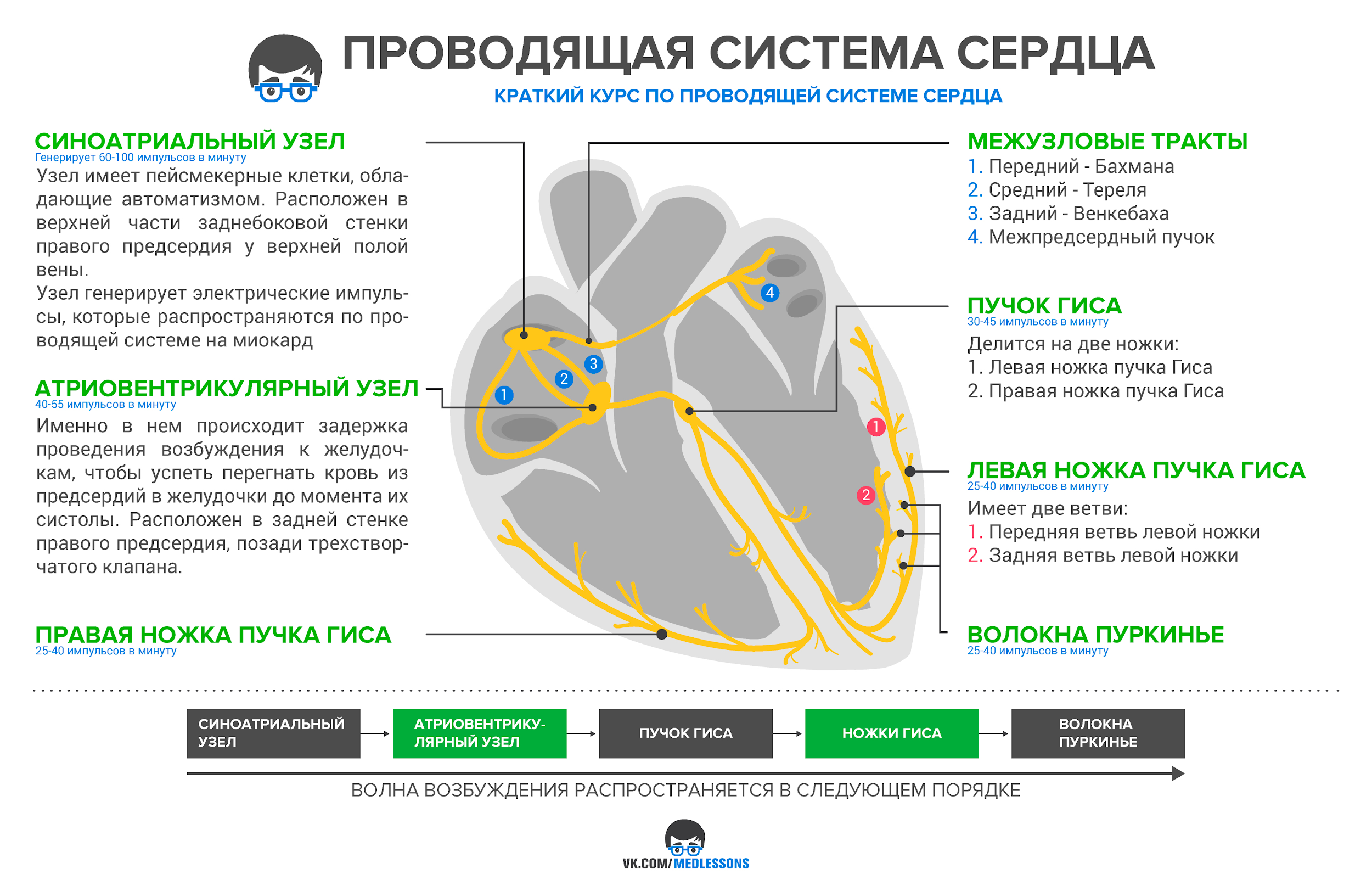
Обеспечивает автоматизм сердца
Благодаря задержке в передаче сигнала, сначала сокращаются предсердия, а потом желудочки
Органы кроветворения и иммуногенеза
1. Органы кроветворения и иммуногенеза: классификация, общие принципы строения и функции. Унитарная система кроветворения А.А. Максимова и ее современная трактовка. Понятие о стволовых кроветворных клетках. Эмбриональное и постэмбриональное кроветворение.
Система кроветворения – это функционально связанные между собой специализированные органы, кровь, лимфа и лимфоидная ткань, ассоциированная со слизистыми + клетки (клетки, макрофаги и антигенпредставляющие клетки), которые находятся в других тканях
Кроветворение подразделяют на лимфоидное (дозревание и работа Т-лимфоцитов) и миелоидное (вся кровь, все стадии В-лимфоцитов и ранние стадии Т-лимфоцитов)
Органы кроветворения делятся на центральные (по сути, отвечают за миелоидное кроветворение) и периферические (больше по лимфоидному - завершают дифференцировку Т-лимфоцитов и размножают оба вида лимфоцитов при необходимости).
Строение органов кроветворения, их компоненты:
А) Стромальный. Он представлен
1) ретикулярной тканью — в красном костном мозгу, лимфоузлах и селезенке; (про неё упоминалось в разделе с соединительной тканью)
2) рыхлой соединительной тканью — в лимфатических образованиях слизистых оболочек;
3) эпителиальной (ретикулоэпителиальной) тканью — в тимусе.
Б) Второй компонент — гемальный. Это гемопоэтические (кроветворные) клетки на разных стадиях созревания. Они находятся в тесной связи с элементами стромального компонента, образующими микроокружение.
В) Макрофаги и клетки их ряда.
Г) Специфические кровеносные и (в ряде органов) лимфатические сосуды.
Унитарная теория кроветворения:
У всех клеточных популяций крови есть общий предшественник, из которого постепенно дифференцируется любой из элементов кровяного русла.
ГСК (гемопоэтическая стволовая клетка) = СКК (стволовая клетка крови)
По числу разных видов форменных элементов крови существует шесть направлений миелопоэза (образование эритроцитов, моноцитов, тромбоцитов и трех видов гранулоцитов) и не менее двух направлений лимфопоэза. В каждом из этих путей дифференцировки различают шесть классов клеток: 1) ГСК (СКК, стволовые клетки крови), 2) полустволовые клетки, 3) унипотентные клетки, 4) бласты, 5) созревающие клетки, 6) зрелые клетки.
Эти клетки делятся относительно редко (раз в 10–50 суток); в основном же они находятся в G0-периоде. Поэтому их доля в кроветворных органах очень мала. Ряд веществ (интерлейкин-3, колониестимулирующие факторы) активируют деления ГСК. В этих делениях проявляется ключевое свойство стволовых клеток — способность к самоподдержанию. Это значит, что при делениях часть дочерних клеток (не менее 50 %) полностью идентична материнским (что поддерживает численность популяции ГСК), и лишь другая часть подвергается дифференцировке (превращается в клетки последующих классов). ГСК являются полипотентными: могут давать начало всем форменным элементам крови. На первом этапе дифференцировки ГСК образуются полустволовые клетки двух видов: предшественники миелопоэза и предшественники лимфопоэза.
У зародыша по мере его развития локализация кроветворения последовательно меняется. В связи с этим выделяют три этапа эмбрионального гемоцитопоэза: мезобластический, печеночный и медуллярный.
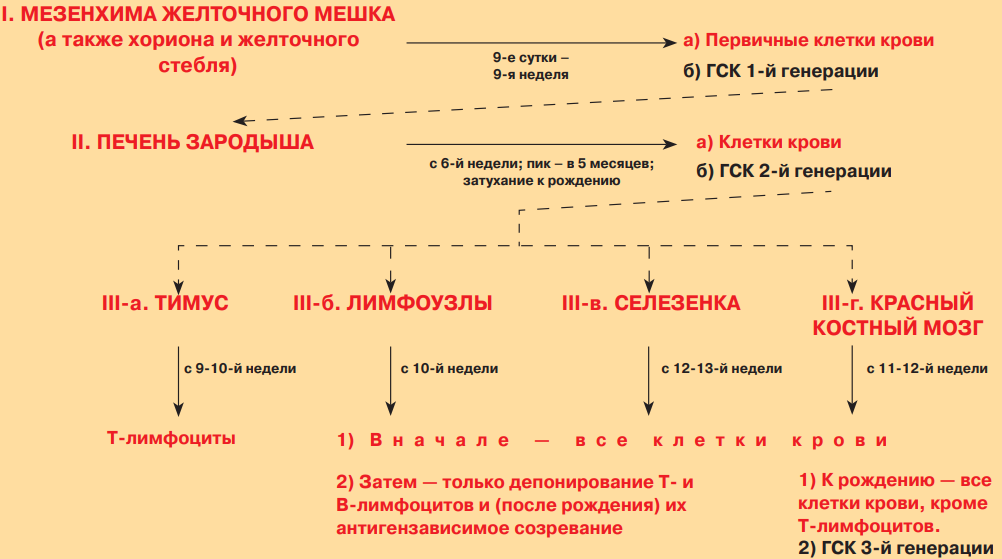
2. Органы кроветворения и иммуногенеза: классификация, общие принципы строения и функции. Строение красного костного мозга, особенности кровоснабжения. Гемопоэтические клетки и диффероны гемопоэза. Регуляция гемопоэза в постэмбриональный период.
Особенности строения красного костного мозга: гемопоэтические клетки располагаются в красном костном мозгу островками. Преобладают же в островках клетки класса V гемопоэза. Кровеносные сосуды подходят к красному костному мозгу со стороны костного вещества. Капиллярная сеть этого вещества продолжается в капиллярную сеть красного костного мозга. В последнем же одни капилляры остаются узкими (6–20 мкм) и выполняют трофическую роль. Другие капилляры переходят в широкие тонкостенные синусы (200–500 нм), имеющие щелевидные поры в эндотелии и базальной мембране. Эти сосуды называют синусными капиллярами, или венозными синусами. Они обеспечивают избирательный выход в кровь только достаточно зрелых форменных элементов (определение зрелости производится, вероятно, по наличию определенных поверхностных гликопротеинов). В самих синусах происходит дозревание эритроцитов. Кроме того, эндотелиоциты синусов, видимо, синтезируют факторы, стимулирующие кроветворение
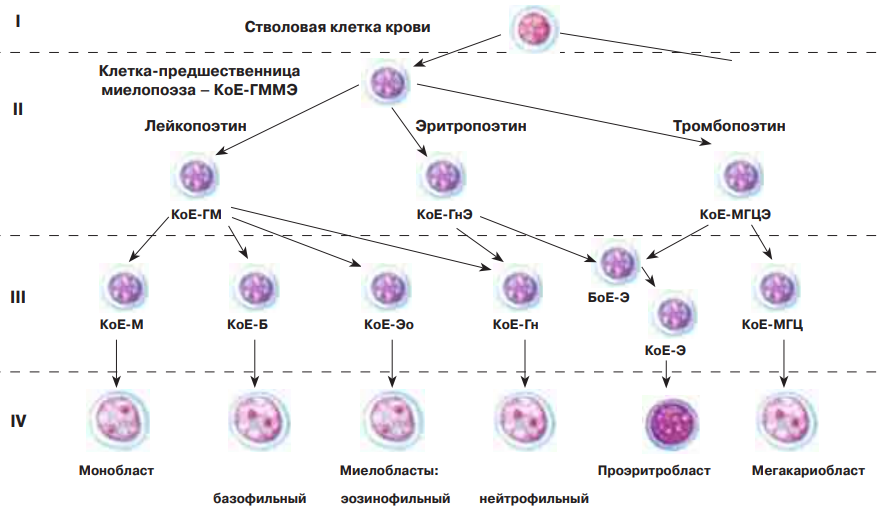
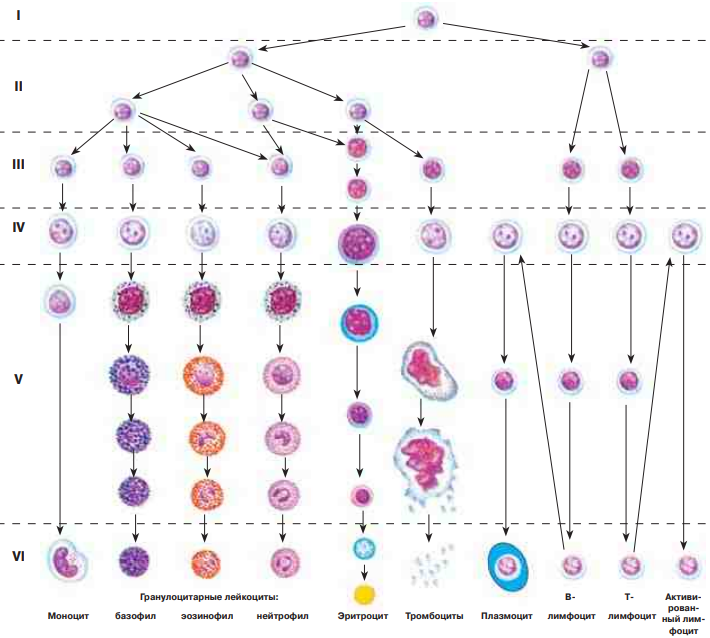
Эритроцитарный дифферон:
1) СКК
2) предшественница миелопоэза
3) колониеобразующая клетка эритроцитопоэза
4) эритробласт
5) проэритроцбласт, базофильный эритроцбласт, полихроматофильный эритроцбласт, оксифильный эритроцбласт, ретикулоцит
6) ретикулоцит и эритроцит
Гранулоцитарный дифферон:
1) СКК
2) предшественница миелопоэза
3) колониеобразующая клетка + название гранулоцита (эозинофильная, базофильная, нейтрофильная)
4) миелобласт
5) Промиелоцит, миелоцит, метамиелоцит, палочкоядерный (эозинофил, базофил, нейтрофил)
6) Сегментоядерный (эозинофил, базофил, нейтрофил)
Моноцитарный дифферон:
1) СКК
2) Предшественница миелопоэза
3) колониеобразующая моноцитарная клетка
4) монобласт
5) промоноцит
6) моноцит
Трмбоцитарный дифферон:
1) СКК
2) предшественница миелопоэза
3) колониеобразующая мегакариоцитарная клетка
4) мегакариобласт
5) промегакариоцит, мегакариоцит
6) тромбоцит
Лимфоцитарный дифферон:
1) СКК
2) предшественница лимфопоэза
3) пре-В(Т)-клетка
4) В(Т)-лимфобласт
5) В(Т)-пролимфоцит
6) В(Т)-лимфоцит
Регуляция гемопоэза:
«+» Колониестимулирующие факторы: мультипотенциальный КСФ, эритропоэтин, тромбопоэтин, КСФ гранулоцитов, тимопоэтины и др. Эти факторы действуют преимущественно на клетки ранних стадий гемопоэза, определяя направление их дифференцировки. Интерлейкины: ИЛ-1, ИЛ-2, ... ИЛ-7. Последние влияют, в основном, на относительно дифференцированные клетки (определяя их активность, пролиферативную способность и т.д.).
« – » Среди специфических ингибиторов гемопоэза наиболее известны кейлоны. Они вырабатываются зрелыми клетками крови и угнетают кроветворение. Таким образом, здесь имеет место регуляция по принципу отрицательной обратной связи. А среди относительно неспецифических ингибиторов можно назвать, в частности, т.н. тканевые гормоны – простагландины, интерферон, а также истинные гормоны – глюкокортикоиды. Последние особенно сильно угнетают лимфопоэз, уменьшая объем лимфоидной ткани.
3. Органы кроветворения и иммуногенеза: классификация, общие принципы строения и функции. Тимус: строение, участие тимуса в формировании иммунной системы, эндокринная функция, возрастная и акцидентальная инволюция. Характеристика клеток «микроокружения» для тимоцитов коркового и мозгового вещества.
Тимус отличается от прочих кроветворных органов тем, что его строма имеет эпителиальную природу: происходит из эпителия передней части первичной кишки. Гемальный компонент тимуса происходит из предшественников Т-клеток— унипотентных клеток (класс III), мигрирующих в тимус из красного костного мозга. С поверхности тимус покрыт капсулой из плотной волокнистой соединительной ткани. От нее отходят перегородки, которые тоже являются соединительнотканными и разделяют тимус на дольки. В каждой дольке различают корковое и мозговое вещество. Корковое вещество составляет периферическую часть дольки и является на препарате более темным, поскольку густо заселено лимфоцитами. Мозговое вещество — находится в средней части дольки и является более светлым — из-за меньшего содержания лимфоцитов. Как и прочие кроветворные органы, и корковое, и мозговое вещество долек тимуса включает четыре компонента: созревающие или зрелые Т-клетки, эпителиальная строма, клетки макрофагического ряда, особые кровеносные, а также лимфатические сосуды. В подкапсулярной области коркового вещества дольки находятся Т-лимфобласты — клетки класса IV, которые образуются из унипотентных предшественников Т-лимфоцитов. По виду Т-лимфобласты более крупные и светлые, чем зрелые лимфоциты. При этом Т-лимфобласты несколько раз делятся. Основная же часть коркового вещества занята клетками класса V — созревающими Т-лимфоцитами. Эти клетки проходят через 2 процесса: вначале в клетках осуществляется реаранжировка генов — формирование полного гена, кодирующего Т-клеточный рецептор (TCR) с той или иной иммуноспецифичностью, затем проходят два тура селекции клеток по природе их TCR. В частности, второй тур (отрицательной селекции) совершается уже на границе коркового и мозгового вещества и приводит к отбраковке и уничтожению тех Т-клеток, чьи TCR способны взаимодействовать с собственными антигенными детерминантами организма. Внутри долек строму образуют ретикулоэпителиальные, клетки. Они участвуют в положительной селекции Т-клеток, кроме того, выполняют опорную, секреторную и трофическую функции. Выделяют 3 вида ретикулоэпителиоцитов: опорные, секреторные (звёздчатые) (продуцируют тимопоэтин и тимозин => стимулируют пролиферацию Т-клеток в тимусе и других периферических органах), клетки-няньки (трофическая функция, в цитоплазме есть глубокие инвагинации, в которых могут одновременно развиваться около 10 Т-клеток). В корковом веществе помимо типичных макрофагов встречаются дендритные и интердигитирующие клетки, которые в ходе отрицательной селекции Т-клеток представляют им на своей поверхности антигены ГКГ-I и ГКГ-II и связанные с ними пептиды — собственные антигенные детерминанты организма. В мозговом веществе есть слоистые эпителиальные тельца (тельца Гассаля), функция которых неизвестна.
Инволюция тимуса: после 20 лет начинается возрастная инволюция (уменьшение массы) тимуса. Иногда может иметь место и т. н. акцидентальная (быстрая) инволюция — в ответ на стрессовые ситуации, под влиянием гормонов стресса — глюкокортикоидов.
Лимфопоэз включает два этапа: – антигеннезависимое созревание лимфоцитов, происходящее в центральных органах кроветворения, – и антигензависимую дифференцировку, которая следует после встречи лимфоцитов с антигенами и осуществляется в периферической лимфоидной ткани. У взрослого человека антигеннезависимое созревание постепенно затухает и основным источником пополнения популяции лимфоцитов в организме становится антигензависимый лимфопоэз.
Ключевые события антигеннезависимого лимфопоэза: на данном этапе как раз и происходит формирование у лимфоцитов иммуноспецифичности. Это достигается с помощью ряда уникальных процессов, из которых наиболее важными являются следующие:
а) Ключевой процесс – образование генов Ig-подобных мембранных рецепторов – путем перестройки (реаранжировки) соответствующей геномной области. Причем результат этой перестройки в разных клетках различен.
б) Затем следует отбор (селекция) появившихся клонов лимфоцитов: Вначале (в первом туре) отбор происходит по способности клеток взаимодействовать с антигенами вообще: клетки с указанной способностью сохраняются, что обозначается как положительная селекция (В качестве «инспекторов», проверяющих способность созревающих Т-клеток к взаимодействию, выступают эпителиальные клетки стромы тимуса.). Во втором туре отбор идет по способности взаимодействовать с собственными антигенами организма: клетки с указанной способностью уничтожаются – отрицательная селекция (Они вступают во взаимодействие с клетками макрофагического ряда — дендритными и интердигитирующими, которые, как полагают, представляют Т-клеткам антигенные детерминанты (пептиды), встречающиеся в составе собственных белков организма).
Чем вызвана необходимость в реаранжировке генов? Дело в том, что геном человека (да и любого другого организма) не может содержать гены антител (иммуноглобулинов) сразу против всех возможных антигенов. Это потребовало бы на порядок увеличить его размеры: вместо 46 хромосом понадобилось бы, например, 460 хромосом. Поэтому в хромосомах любой клетки организма (в т.ч. гемопоэтической стволовой клетки, ГСК) содержатся не длинные гены целых пептидных цепей всевозможных иммуноглобулинов (Ig), а короткие гены нескольких пептидных участков, из которых состоит цепь Ig. Причем, для каждого такого участка имеется до нескольких сотен аллелей (вариантов) его короткого гена. В очень упрощенном виде это можно представить так. Будем считать, что полная цепь Ig состоит из четырех участков (a, b, c, d), и геном содержит 300 различных аллелей гена A, 20 аллелей гена B, 200 аллелей гена C и 10 аллелей гена D. Вместе это составляет всего 530 коротких генов — вместо миллиардов длинных генов полных цепей Ig.
Суть реаранжировки генов - в ходе лимфопоэза ферменты рекомбиназы «выхватывают» из каждого аллельного множества по одному произвольно выбранному короткому гену и «сшивают» «избранников» в один (в Т-клетках) или в два (в В-клетках) полных гена Ig. В результате, в каждой клетке образуются гены лишь для одного вида Ig (состоящего из цепей одного или двух видов), но общее количество вариантов полных генов, а, следовательно, и самих Ig — а, значит, и лимфоцитов с разной иммунной специфичностью, — получается весьма большим.
Сохранившиеся клетки размножаются, так что из каждой клетки образуется клон — совокупность лимфоцитов (пока еще незрелых) с одинаковой иммунной специфичностью. По разным оценкам, число таких клонов (отдельно для В- и отдельно для Т-лимфоцитов) варьирует от 107 до 1010. Затем новые гены (продукты реаранжировки) начинают работать, и продукты их экспрессии появляются на поверхности лимфоидных клеток: в случае В-клеток это иммуноглобулины классов М (IgM) и D (IgD), выступающие в качестве BCR, а в случае Т-клеток — более просто устроенные Ig-подобные белки, TCR. Каждый такой рецептор способен взаимодействовать лишь с одним определенным антигеном — таким, чья структура комплементарна рецептору.
В итоге подавляющая доля (до 90 %) созревающих в тимусе Т-клеток погибает. Причем гибель клеток происходит не только из-за неудачной реаранжировки генов и последующей селекции, но и из-за токсического действия продуктов распада ДНК (метаболитов пурина), которые попадают в межклеточную среду тимуса при массовом разрушении тимоцитов.
По окончании антигеннезависимого созревания В-лимфоциты (из костного мозга) и Т-лимфоциты (из тимуса) перемещаются в периферические лимфоидные органы (лимфоузлы, селезенку, лимфатические узелки слизистых оболочек), а также в кожу. При этом в периферических лимфоидных органах имеются – В-зоны — участки преимущественной локализации В-клеток и продуктов их дифференцировки, а также – Т-зоны — места сосредоточения Т-клеток. Клетки, еще не встречавшиеся с антигеном, называются наивными, или виргильными (девственными).