

- •Contents
- •Preface to the first edition
- •Flagella
- •Cell walls and mucilages
- •Plastids
- •Mitochondria and peroxisomes
- •Division of chloroplasts and mitochondria
- •Storage products
- •Contractile vacuoles
- •Nutrition
- •Gene sequencing and algal systematics
- •Classification
- •Algae and the fossil record
- •REFERENCES
- •CYANOPHYCEAE
- •Morphology
- •Cell wall and gliding
- •Pili and twitching
- •Sheaths
- •Protoplasmic structure
- •Gas vacuoles
- •Pigments and photosynthesis
- •Akinetes
- •Heterocysts
- •Nitrogen fixation
- •Asexual reproduction
- •Growth and metabolism
- •Lack of feedback control of enzyme biosynthesis
- •Symbiosis
- •Extracellular associations
- •Ecology of cyanobacteria
- •Freshwater environment
- •Terrestrial environment
- •Adaption to silting and salinity
- •Cyanotoxins
- •Cyanobacteria and the quality of drinking water
- •Utilization of cyanobacteria as food
- •Cyanophages
- •Secretion of antibiotics and siderophores
- •Calcium carbonate deposition and fossil record
- •Chroococcales
- •Classification
- •Oscillatoriales
- •Nostocales
- •REFERENCES
- •REFERENCES
- •REFERENCES
- •RHODOPHYCEAE
- •Cell structure
- •Cell walls
- •Chloroplasts and storage products
- •Pit connections
- •Calcification
- •Secretory cells
- •Iridescence
- •Epiphytes and parasites
- •Defense mechanisms of the red algae
- •Commercial utilization of red algal mucilages
- •Reproductive structures
- •Carpogonium
- •Spermatium
- •Fertilization
- •Meiosporangia and meiospores
- •Asexual spores
- •Spore motility
- •Classification
- •Cyanidiales
- •Porphyridiales
- •Bangiales
- •Acrochaetiales
- •Batrachospermales
- •Nemaliales
- •Corallinales
- •Gelidiales
- •Gracilariales
- •Ceramiales
- •REFERENCES
- •Cell structure
- •Phototaxis and eyespots
- •Asexual reproduction
- •Sexual reproduction
- •Classification
- •Position of flagella in cells
- •Flagellar roots
- •Multilayered structure
- •Occurrence of scales or a wall on the motile cells
- •Cell division
- •Superoxide dismutase
- •Prasinophyceae
- •Charophyceae
- •Classification
- •Klebsormidiales
- •Zygnematales
- •Coleochaetales
- •Charales
- •Ulvophyceae
- •Classification
- •Ulotrichales
- •Ulvales
- •Cladophorales
- •Dasycladales
- •Caulerpales
- •Siphonocladales
- •Chlorophyceae
- •Classification
- •Volvocales
- •Tetrasporales
- •Prasiolales
- •Chlorellales
- •Trebouxiales
- •Sphaeropleales
- •Chlorosarcinales
- •Chaetophorales
- •Oedogoniales
- •REFERENCES
- •REFERENCES
- •EUGLENOPHYCEAE
- •Nucleus and nuclear division
- •Eyespot, paraflagellar swelling, and phototaxis
- •Muciferous bodies and extracellular structures
- •Chloroplasts and storage products
- •Nutrition
- •Classification
- •Heteronematales
- •Eutreptiales
- •Euglenales
- •REFERENCES
- •DINOPHYCEAE
- •Cell structure
- •Theca
- •Scales
- •Flagella
- •Pusule
- •Chloroplasts and pigments
- •Phototaxis and eyespots
- •Nucleus
- •Projectiles
- •Accumulation body
- •Resting spores or cysts or hypnospores and fossil Dinophyceae
- •Toxins
- •Dinoflagellates and oil and coal deposits
- •Bioluminescence
- •Rhythms
- •Heterotrophic dinoflagellates
- •Direct engulfment of prey
- •Peduncle feeding
- •Symbiotic dinoflagellates
- •Classification
- •Prorocentrales
- •Dinophysiales
- •Peridiniales
- •Gymnodiniales
- •REFERENCES
- •REFERENCES
- •Chlorarachniophyta
- •REFERENCES
- •CRYPTOPHYCEAE
- •Cell structure
- •Ecology
- •Symbiotic associations
- •Classification
- •Goniomonadales
- •Cryptomonadales
- •Chroomonadales
- •REFERENCES
- •CHRYSOPHYCEAE
- •Cell structure
- •Flagella and eyespot
- •Internal organelles
- •Extracellular deposits
- •Statospores
- •Nutrition
- •Ecology
- •Classification
- •Chromulinales
- •Parmales
- •Chrysomeridales
- •REFERENCES
- •SYNUROPHYCEAE
- •Classification
- •REFERENCES
- •EUSTIGMATOPHYCEAE
- •REFERENCES
- •PINGUIOPHYCEAE
- •REFERENCES
- •DICTYOCHOPHYCEAE
- •Classification
- •Rhizochromulinales
- •Pedinellales
- •Dictyocales
- •REFERENCES
- •PELAGOPHYCEAE
- •REFERENCES
- •BOLIDOPHYCEAE
- •REFERENCE
- •BACILLARIOPHYCEAE
- •Cell structure
- •Cell wall
- •Cell division and the formation of the new wall
- •Extracellular mucilage, biolfouling, and gliding
- •Motility
- •Plastids and storage products
- •Resting spores and resting cells
- •Auxospores
- •Rhythmic phenomena
- •Physiology
- •Chemical defense against predation
- •Ecology
- •Marine environment
- •Freshwater environment
- •Fossil diatoms
- •Classification
- •Biddulphiales
- •Bacillariales
- •REFERENCES
- •RAPHIDOPHYCEAE
- •REFERENCES
- •XANTHOPHYCEAE
- •Cell structure
- •Cell wall
- •Chloroplasts and food reserves
- •Asexual reproduction
- •Sexual reproduction
- •Mischococcales
- •Tribonematales
- •Botrydiales
- •Vaucheriales
- •REFERENCES
- •PHAEOTHAMNIOPHYCEAE
- •REFERENCES
- •PHAEOPHYCEAE
- •Cell structure
- •Cell walls
- •Flagella and eyespot
- •Chloroplasts and photosynthesis
- •Phlorotannins and physodes
- •Life history
- •Classification
- •Dictyotales
- •Sphacelariales
- •Cutleriales
- •Desmarestiales
- •Ectocarpales
- •Laminariales
- •Fucales
- •REFERENCES
- •PRYMNESIOPHYCEAE
- •Cell structure
- •Flagella
- •Haptonema
- •Chloroplasts
- •Other cytoplasmic structures
- •Scales and coccoliths
- •Toxins
- •Classification
- •Prymnesiales
- •Pavlovales
- •REFERENCES
- •Toxic algae
- •Toxic algae and the end-Permian extinction
- •Cooling of the Earth, cloud condensation nuclei, and DMSP
- •Chemical defense mechanisms of algae
- •The Antarctic and Southern Ocean
- •The grand experiment
- •Antarctic lakes as a model for life on the planet Mars or Jupiter’s moon Europa
- •Ultraviolet radiation, the ozone hole, and sunscreens produced by algae
- •Hydrogen fuel cells and hydrogen gas production by algae
- •REFERENCES
- •Glossary
- •Index

APICOMPLEXA 313
REFERENCES
Fast, N. M., Kissinger, J. C., Roos, D. S., and Keeling, P. J. (2001). Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol. Biol. Evol. 18:418–26.
Foth, B. J., and McFadden, G. I. (2003). The apicoplast: a plastid in Plasmodium falciparum and other apicomplexan parasites. Int. Rev. Cytol. 224:57–110.
Lee, R. E., and Kugrens, P. (1991). Katablepharis ovalis, a colorless flagellate with interesting cytological characteristics. J. Phycol. 27:505–15.
Lee, R. E., Kugrens, P., and Mylnikov, A. P. (1991). Feeding apparatus of the colorless flagellate Katablepharis (Cryptophyceae). J. Phycol. 27:725–33.
Ralph, S. A., van Dooren, G. G., Waller, R. F., et al. (2004). Metabolic maps and functions of the
Plasmodium falciparum apicoplast. Nat. Rev./Microbio. 2:203–16.
Sam-Yellowe, T. Y. (1996). Rhoptry organelles of the Apicomplexa: their role in host cell invasion and intracellular survival. Parasitol. Today 12:308–16.
Wilson, R. J. (1993). Plastids better red than dead. Nature 366:638.
Wilson, R. J., Williamson, D. H., and Preiser, P. (1994). Malaria and other apicomplexans: the “plant” connection. Infect. Agents Dis. 3:29–37.

Part V
Evolution of two membranes of chloroplast endoplasmic reticulum and the Chlorarachniophyta
Algae with two membranes of chloroplast endoplasmic reticulum (chloroplast E.R.) have the inner membrane of chloroplast E.R. surrounding the chloroplast envelope. The outer membrane of chloroplast E.R. is continuous with the outer membrane of the nuclear envelope and has ribosomes on the outer surface (Fig. V.1).
The algae with two membranes of chloroplast E.R. evolved by a secondary endosymbiosis (Fig. V.1) (Lee, 1977) when a phagocytic protozoan took up a eukaryotic photosynthetic alga into a food vesicle. Instead of being phagocytosed by the protozoan, the photosynthetic alga became established as an endosymbiont within the food vesicle of the protozoan. The endosymbiotic photosynthetic alga benefited from the acidic environment in the food vesicle that kept much of the inorganic carbon in the form of carbon dioxide, the form needed by ribulose bisphosphate/ carboxylase for carbon fixation (see Part IV for further explanation). The host benefited by receiving some of the photosynthate from the endosymbiotic alga. The food vesicle membrane eventually fused with the endoplasmic reticulum of the host protozoan, resulting in ribosomes on the outer surface of this membrane, which became the outer membrane of the chloroplast E.R. Through evolution, ATP production and other functions of the endosymbiont’s mitochondrion were taken over by the mitochondria of the protozoan host, and the mitochondria of the endosymbiont were lost. The host nucleus also took over some of the genetic control of the endosymbiont, with a reduction in the size and function of the nucleus of the endosymbiont. The resulting cytology is characteristic of the extant algae in the Chlorarachniophyta and Cryptophyta, which have a nucleomorph representing the degraded endosymbiotic nucleus, as well as storage product produced in what remains of the endosymbiont cytoplasm.
The type of chloroplast E.R. that exists in the Heterokontophyta and the Prymnesiophyta resulted from further reduction. The nucleomorph
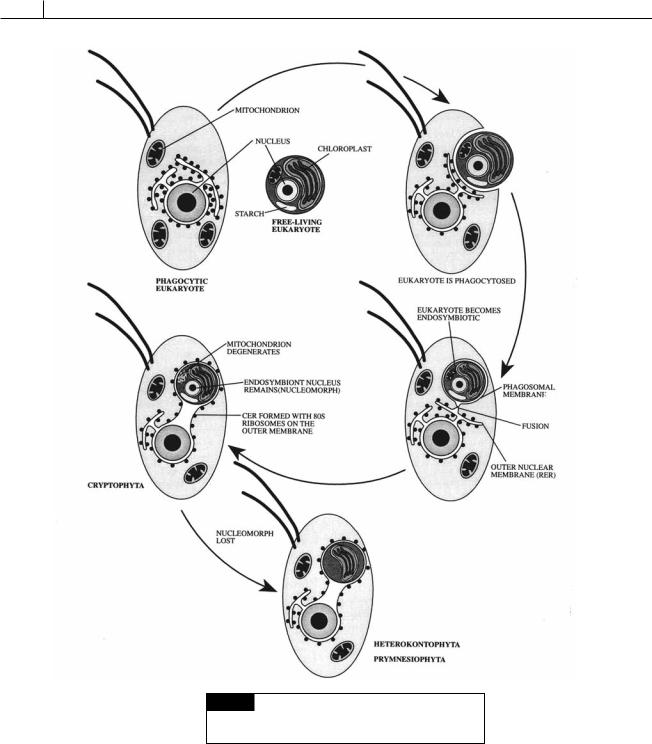
316
Fig. V.1 The sequence of events that led to the evolution
of algae with two membranes of chloroplast endoplasmic
reticulum. (Drawing by Brec Clay.)
was completely lost and storage product formation was taken over by the host. The resulting cell had two membranes of chloroplast envelope surrounding the chloroplast. Outside of this was the inner membrane of chloroplast E.R. that was the remains of the plasma membrane of the endosymbiont. Outside of this was the outer membrane of