

150 EVOLUTION OF THE CHLOROPLAST
oxygen radicals (O2 ) and other toxic oxygen species (OH·, singlet oxygen) and convert them to less toxic moieties.
There are three forms of superoxide dismutases, named for the metal in their reaction centers: iron SOD (FeSOD), manganese SOD (MnSOD), and copper-zinc SOD (Cu/Zn SOD). The amino acid sequences and three-dimensional protein structure of FeSOD and MnSOD are very similar, suggesting a close evolutionary relationship, whereas those of Cu/Zn SOD are quite different, indicating very little, if any, evolutionary relationship to the other two (deJesus et al., 1989).
FeSOD and MnSOD occur in all classes of green algae and most land plants. Cu/Zn SOD, however, occurs only in the Charophyceae and land plants, reinforcing the concept that organisms similar to those in the Charophyceae evolved into land plants.
Cu/Zn SOD is the only superoxide dismutase that occurs in the cytosol. MnSOD occurs in the mitochondrion while FeSOD occurs in the chloroplast. DeJesus et al. (1989) argue that green algae evolving to land plants would need superoxide dismutase in the cytosol to counteract oxygen diffusing in from the atmosphere, which would produce damaging radicals in the cytosol. The charophycean lineage of green algae is the only one to have superoxide dismutase in the cytosol and is a major reason for the successful colonization of the land by descendants of the Charophyceae. The other lineages of green algae were unable to make the transition to the land because they had superoxide dismutase restricted to mitochondria, chloroplasts, and peroxisomes and were not able to counteract the production of damaging radicals formed by oxygen diffusing in from the atmosphere.
Prasinophyceae
The Prasinophyceae (Micromonadophyceae) contain primitive green flagellates with scales composed of acidic polysaccharides (Wustman et al., 2004). The class is a catch-all for those algae that “exhibit primitive characteristics for many of the features used to define other classes” (Mattox and Stewart, 1984; Fawley et al., 2000).
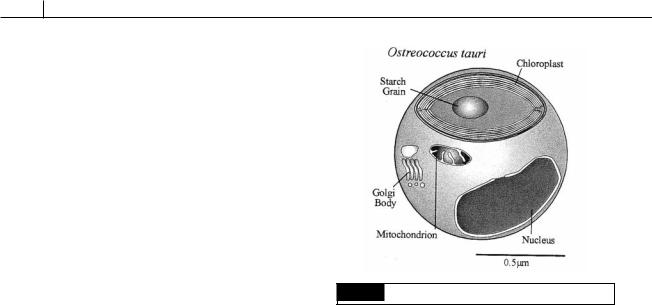
Fig. 5.9 The general organization of Ostreococcus tauri.
Ostreococcus tauri (Fig. 5.9) is less than 1 m in diameter and is the smallest eukaryotic alga known. The alga has a relatively large nucleus, a single chloroplast with a starch granule, a mitochondrion, and a Golgi apparatus. Unlike other algae in the group, there are no scales outside the cell wall (Courties et al., 1998).
Pyramimonas obovata has heart-shaped (cordate) cells, with four flagella arising from an anterior flat-bottomed depression (Fig. 5.10). There are three different layers of scales on the body and two layers on the flagellum (Belcher et al., 1974). The scales are formed in dictyosomes, from which they are transported to the scale reservoir and then to the surface of the cell in vesicles.
Species of Pyramimonas have adapted to tidepools by means of a settling rhythm. Cells in tidepools anticipate the incoming tide and move down into the sand where they attach to sand grains by their flagella, before the first waves cover the tide pool. After the tide pool is exposed by the receding tide, the Pyramimonas cells move upward into the tide pool. The settling is controlled by an endogenous circadian oscillator and will continue in the laboratory for a few days after the cells are removed from the wild (Griffin and Aken, 1993).
Pterosperma is a genus that is closely related to
Pyramimonas. Pterosperma produces walled cysts called phycomata (Fig. 5.11) that are similar to microfossils such as Tasmanites and Cymatiosphaera that have been reported from the Precambrian up to the Holocene (Inouye et al., 1990).
CHLOROPHYTA 151
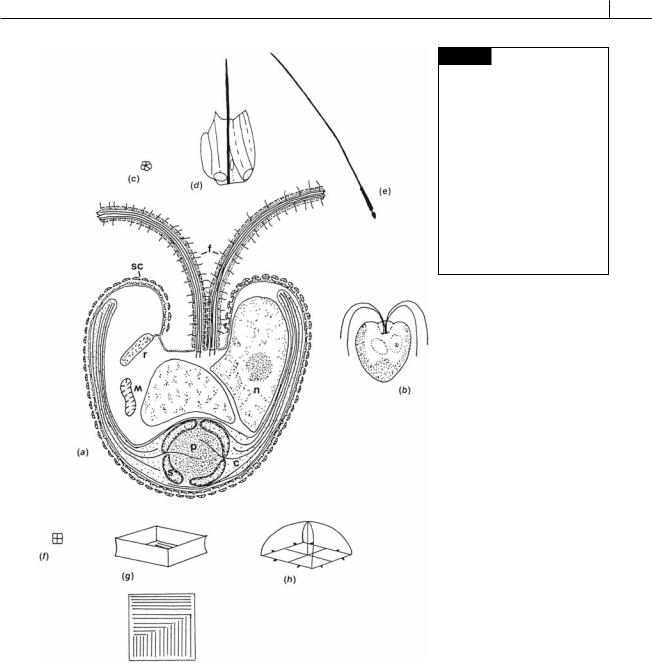
Fig. 5.10 Pyramimonas obovata. (a) Semidiagrammatic drawing of a cell showing the organelles, two of the four flagella, and the layers of scales on the body and the flagella. (b) Whole cell. (c) Inner flagella scale. (d) Outer flagellar scale.
(e) Flagellar hair. (f) Inner body scale. (g) Intermediate body scale, top and side view. (h) Outer body scale. (c) Chloroplast; (f) flagellum;
(m) mitochondrion; (n) nucleus;
(p) pyrenoid; (r) scale reservoir;
(s) starch; (sc) scales. (After Belcher et al., 1974.)
Tetraselmis (Fig. 5.12) is commonly found in marine waters. The cells are oval-shaped and surrounded by a theca that is formed by the coalescence of many small stellate scales that are produced in the Golgi apparatus. Four flagella are inserted in an apical pit in the cell. The flagella are covered with hairs and scales, and the flagella emerge from an opening in the theca. There is a cup-shaped chloroplast with a basal pyrenoid. A
nucleus occurs in the center of the cell, and Golgi bodies are found anterior to the nucleus.
Tetraselmis produces stellate scales in the Golgi apparatus which fuse extracellularly to yield the theca (Manton and Parke, 1960). The theca is composed of neutral and acidic polysaccharides associated with certain amino acids (Becker et al., 1996), similar to the glycoprotein walls in volvocean flagellates (those green algae in the
152 EVOLUTION OF THE CHLOROPLAST
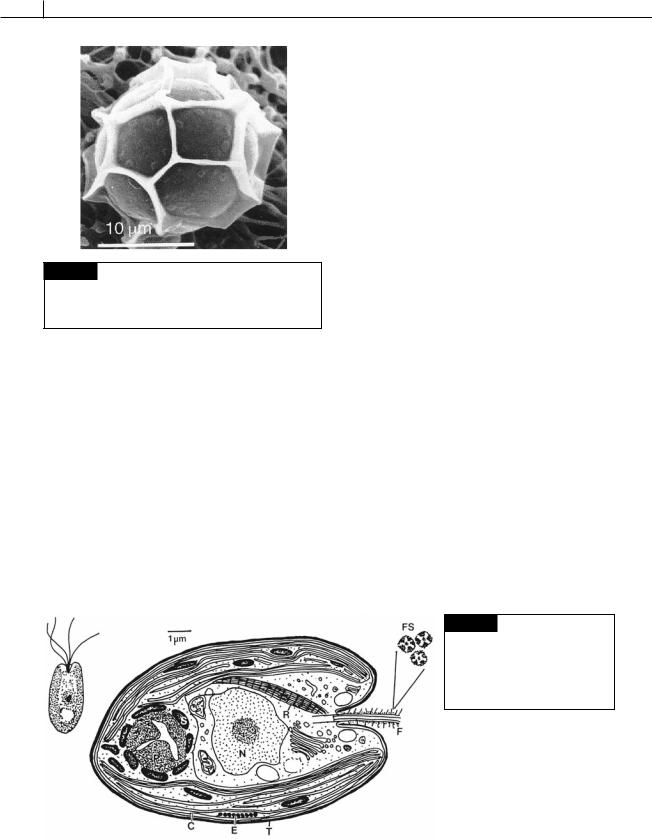
Fig. 5.11 A scanning electron micrograph of a phycoma of
Pterosperma cristatum. Square, pentagonal or hexagonal compartments are formed by wing-like protrusions of the cell wall. (From Inouye et al., 1990.)
Volvocales) (Roberts, 1974) but unlike the cellulose cell walls in the Charophyceae. The typical volvocean flagellates and eventually the rest of the higher Chlorophyceae probably arose from a flagellate such as Tetraselmis or Mantionella (Fig. 5.54) (Domozych et al., 1981).
The genus Tetraselmis was previously divided into a number of genera including Tetraselmis,
Platymonas, and Prasinocladus before it was recognized that all of the cells were similar. All of the organisms are now recognized as the single genus Tetraselmis with subgenera (Hori et al., 1982a,b):
Subgenus Tetraselmis: spherical pyrenoid into which narrow channels of cytoplasm extend.
Subgenus Prasinocladia: pyrenoid not penetrated by cytoplasm but by a single lobe of the nucleus so that the pyrenoid appears cup-shaped in the microscope.
Algae in the subgenus Prasinocladia grow as dense yellow-green tufts on rocks or shells in small tide-pools near the high-tide mark (Fig. 5.13) (Chihara, 1963). The plants occur on most temperate coasts. In the mature attached vegetative state, the two to four protoplasts are at the apical area of the wall. Protoplasts are released as zoospores by escaping through a break in the apical area of the wall. The quadriflagellate zoospores swim actively for a while, settle, lose their flagella, and secrete a wall. Germination occurs immediately after settling and first involves the secretion of an outer wall. The protoplast elongates with the deposition of an inner wall, causing a break in the outer wall; and, still surrounded by the elongating inner wall, the protoplast secretes additional wall material and positions itself at the top of the resulting tube. The protoplasts can divide into two cells to form new tubes. Any protoplast is capable of forming flagella and becoming a zoospore.
Convoluta is a small flatworm, several millimeters long (Fig. 5.14(a)), that lives on sandy marine shores (Russell and Yonge, 1963). It occurs in large colonies forming green patches on the yellow sand, appearing from beneath the sand immediately after the tide has left it and disappearing just before the tide returns. The green color of the flatworm is due to the presence in the animal of vast numbers of algae. The algae are not present
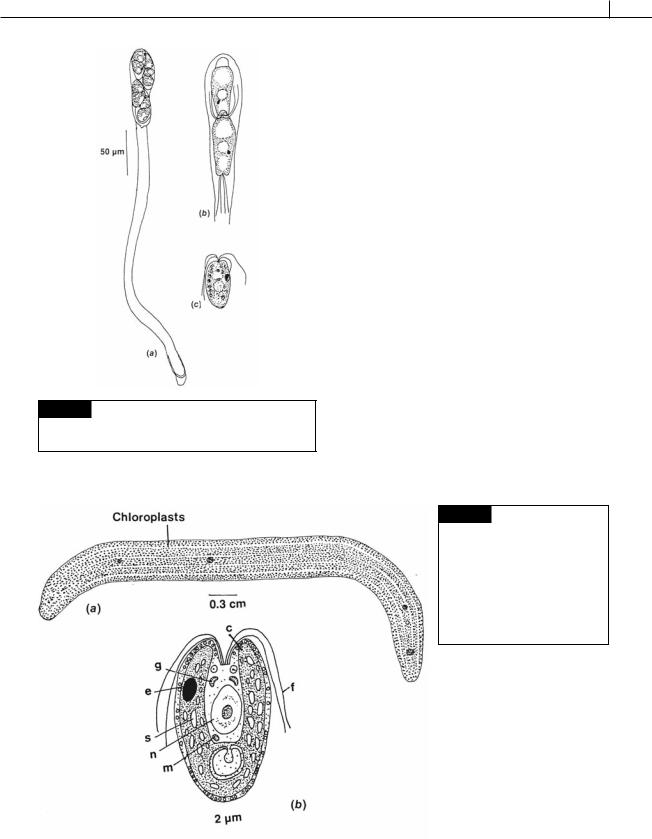
Fig. 5.12 Drawing of a cell of
Tetraselmis. (C) Chloroplast;
(E) eyespot; (F) flagellum; (FS) flagellar scales; (N) nucleus; (P) pyrenoid; (R) rhizoplast (only one of two is shown); (S) starch; (T) theca.
CHLOROPHYTA 153
Fig. 5.13 Tetraselmis subgenus Prasinocladia. (a) Vegetative
cells on a stalk. (b) Zoospores within a stalk. (c) Zoospore.
(After Proskauer, 1950.)
in the egg, but there are algae adhering to the egg case that are ingested by the emerging worms. The alga that infects Convoluta roscoffensis is Tetraselmis
(Fig. 5.14(b)) (Oschman, 1966). These algae lie in the extracellular spaces between adjacent cells of the animal. The infecting algae undergo morphological alterations within the worms, losing their flagella, theca, and eyespot in that order. The Tetraselmis theca does not contain cellulose (Lewin, 1958), a possible explanation for the relative ease with which the worm can dissolve the theca and set up the symbiosis. Upon loss of the theca, the alga assumes an irregularly shaped form, with finger-like processes of the algal cells penetrating between adjacent animal cells. Convoluta flatworms may contain algal cells of either the Tetraselmis or Prasinocladia subgenera, although one animal will usually contain cells of only one subgenus. In the United Kingdom, the flatworms having cells of the Tetraselmis subgenus are longer, are more likely to contain gametes, and lay more egg capsules containing more embryos than those containing cells of the Prasinocladia subgenus (Douglas, 1985). If the Convoluta eggs are not infected by Tetraselmis, they fail to develop properly and soon die. In early life, Convoluta, like other flatworms, feeds on smaller
Fig. 5.14 (a) Convoluta roscoffensis
flatworm with symbiotic green algae. (b) Tetraselmis convolutae, the algal symbiont of Convoluta. (c) Chloroplast; (e) eyespot; (f) flagella;
(g) Golgi body; (m) mitochondrion;
(n) nucleus; (s) starch. ((a) after Russell and Yonge, 1963; (b) after Parke and Manton, 1967.)