

242
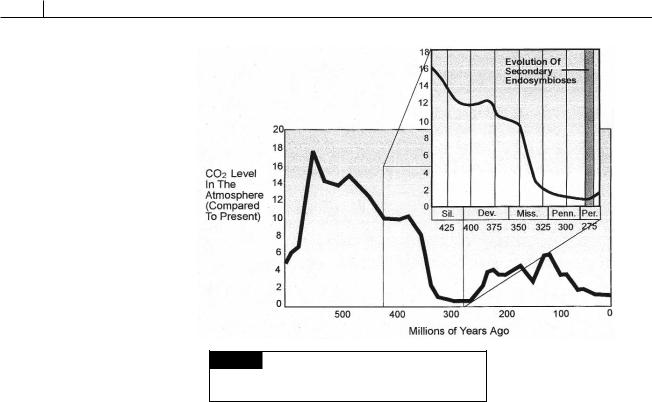
Fig. IV.3 The amount of CO2 in the atmosphere over the past 600 Ma. The time of evolution of secondary endosymbioses around 275 Ma is shown in the insert.
marine environment where at pH 8.2, the dissolved CO2 concentration is particularly low. In such an environment, algae with chloroplast endoplasmic reticulum had a competitive advantage and were selected for in evolution.
On the basis of a molecular clock, Medlin et al. (1997) postulated that the original secondary endosymbiosis occurred between 260 and 285 million years ago (Ma). Atmospheric CO2 levels were at record lows at this time (Fig. IV.3), levels not reached again until Recent times. Such an environment favored algae that were better able to photosynthesize in a low CO2 environment, e.g., those algae with chloroplast endoplasmic reticulum. Thus, the radiation of algae with chloroplast endoplasmic reticulum occurred at a time when the more efficient CO2 mechanism of these algae enabled them to outcompete other algae without chloroplast endoplasmic reticulum.
REFERENCES
Falkowski, P. G., and Raven, J. (1997). Aquatic Photosynthesis. Oxford: Blackwell Science.
Fridlyand, L. E. (1997). Models of CO2 concentrating mechanisms in microalgae taking into account cell and chloroplast structure. BioSystems 44:41–57.
Gibbs, S. P. (1978). The chloroplasts of Euglena may have evolved from symbiotic green algae. Canadian J. Bot. 56:2883–9.
Lee, R. E. (1977). Evolution of algal flagellates with chloroplast endoplasmic reticulum from the ciliates. South African J. of Sci. 73:179–82.

243
Medlin, L. K., Kooistra, W. H. C. F., Gersonde, R., Sims, P. A., and Wellbrock, U. (1997). Is the origin of the diatoms related to the end-Permian mass extinction? Nova Hedwigia 65:1–11.
Raven, J. (1997). CO2 concentrating mechanisms: a direct role for thylakoid lumen acidification? Plant Cell Env. 20:147–54.
Sukenik, A., Tchernov, D., Kaplan, A., Huertas, E. Lubian, L. M., and Livne, A. (1997). Uptake, efflux, and photosynthetic utilization of inorganic carbon by the marine eustigmatophyte Nannochloropsis sp. J. Phycol. 33:969–74.