

БІЛЕТ 30
Амінокислоти, їх фізико-хімічні властивості та класифікація. Будова поліпептидного ланцюга. Первинна структура білків.
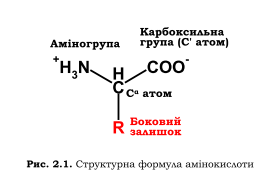
За нейтральних рН аміно- та карбоксильна групи, приєднані до спільного атома карбону (С α), перебувають в іонізованому стані. Саме ці, стандартні для всіх амінокислот, групи (наявність яких і зумовила назву цих сполук) забезпечують утворення полімерної молекули білка. Розрізняються амінокислоти за типом бокового (амінокислотного) залишку. Усього у складі білків зустрічається 20 амінокислот (у дужках - загальноприйняті скорочення, три- та однолітерні): аланін (Ala, A), аргінін (Arg, R), аспарагін (Asn, N), аспарагінова кислота (Asp, D), валін (Val, V), гістидин (His, H), гліцин (Gly, G), глутамін (Gln, Q), глутамінова кислота (Glu, E), ізолейцин (Ile,I), лейцин (Leu, L), лізин (Lys, K), метіонін (Met, M), пролін (Pro, P), серин (Ser, S), тирозин (Tyr, Y), треонін (Thr, T), триптофан (Trp, W), фенілаланін (Phe, P), цистеїн (Cys, C). Два параметри амінокислотних залишків мають особливе значення для структури білка: розмір і здатність взаємодіяти з водою.
За останньою ознакою залишки можна поділити дві групи: полярні й неполярні (гідрофобні). Три гідрофобні залишки маленького розміру, тому їх можна вважати нейтральними щодо спорідненості до води (гідрофобний ефект залежить від розміру поверхні). У глобулярних водорозчинних білках співвідношення між гідрофобними та полярними залишками дорівнює в середньому 50 : 50. Унаслідок гідрофобного ефекту неполярні залишки прагнуть опинитися всередині структури білка, полярні залишаються на поверхні, зберігаючи взаємодії з водою. Це і є головна рушійна сила, яка змушує білковий ланцюг укладатися певним чином у просторі, формуючи компактну структуру - так звану глобулу. Полярні амінокислоти тому так і називаються, що прагнуть взаємодіяти з водою за рахунок водневих зв'язків (містять донорні та акцепторні групи). Серед них є залишки, які мають постійний заряд при рН 7,0: негативний у Asp і Glu (за рахунок карбоксильних груп у складі бокового залишку), позитивний у Lys (аміногрупа) та Arg (гуанідинова група). His також може набувати позитивного заряду в структурі білка в певному мікрооточенні (у розчині він іонізується при рН < 6,5).
Пептидний зв'язок і поліпептидний ланцюг. Карбоксильна група однієї амінокислоти та аміногрупа іншої, втрачаючи молекулу води, утворюють ковалентний зв'язок, який називається пептидним. Група атомів O-C'-N-H називається пептидною групою. Оскільки в утворенні зв'язку між мономерами беруть участь різні хімічні групи, поліпептидний ланцюг має напрям: його кінець, де залишається аміногрупа, називається N-кінцем, протилежний (карбоксильна група) - С-кінцем. Синтезуються поліпептидні ланцюги у напрямку N → С, у цьому ж напрямку представляються за умовчанням амінокислотні послідовності білків.
Дві властивості пептидної групи відіграють ключову роль в утворенні просторової структури білка:
1) Пептидна група є полярною - виступає одночасно донором і акцептором водневих зв'язків. Таким чином, остов поліпептидного ланцюга (пептидні групи та Сα-атоми, що чергуються), від якого відгалужуються полярні та неполярні бокові залишки, є полярним.
2) Одна з пар електронів, що відповідає зв'язку C'-О (зазвичай позначається як подвійний), насправді делокалізована також між атомами C'-N: пептидний зв'язок є «напівподвійним», що
унеможливлює обертання навкруг нього. Для більшості амінокислот реалізується більш вигідна транс-конформація пептидного зв'язку: при- єднані до пептидної групи Сα-атоми «дивляться» в різні боки. Єдине виключення - пептидий зв'язок перед Pro, приблизно 10 % таких зв'язків знаходяться в цис-конформації. Оскільки обертання навкруг пептидного зв'язку є неможливим,
конформація поліпептидного ланцюга може змінюватись тільки за рахунок обертання навкруг суміжних до пептидної групи одинарних зв'язків (обертальні кути φ і ψ), а також обертання навкруг зв'язків у межах бокових залишків (χ 1 , χ 2 ...). Унаслідок стеричних обмежень не всі комбінації кутів обертання φ і ψ є можливими для поліпептидного ланцюга реалізується обертально-ізомерний механізм гнучкості. Дозволені для більшості залишків зони конфірмаційного простору в координатах φ-ψ (діаграма Рамачандрана). Діаграми Рамачандрана мають свої специфічні ознаки для окремих залишків. Особливо це проявляється, якщо ввести третю координату - вільну енергію для кожної пари обертальних кутів. Інакше кажучи, переважна (найбільш імовірна) локальна конформація ланцюга залежить від амінокислотної послідовності.
Ознаки, зчеплені зі статтю.
Спадкування ознак, на які впливає стать, має певні особливості. Зазвичай виділяють три групи таких ознак: ознаки, зчеплені зі статтю; ознаки, обмежені статтю; ознаки, залежні від статі.
Ознаки, зчеплені зі статтю, . це ознаки, гени яких розташовані в статевих хромосомах Х (Z) або Y (W) (ідеться про гени, які зовсім необов’язково мають відношення до статевих ознак). Статеві хромосоми X (Z) і Y (W) суттєво відрізняються одна від одної набором генів. Особина гомогаметної статі може бути як гомо-, так і гетерозиготою за певним геном. Гетерогаметна стать є гемізиготою за більшістю генів: у генотипі існує лише одна копія гена . або в хромосомі Х (Z), або Y (W). Отже,
у гомогаметної статі рецесивна ознака виявляється лише тоді, коли ген перебуває в гомозиготному стані, а в гетерогаметної статі . завжди. Прикладом Х-зчеплених ознак у людини є гемофілія та дальтонізм, а Y-зчеплених . гіпертрихоз (оволосіння вушної раковини) і перетинка між пальцями. Ознаки, зчеплені з Х-хромосомю, спадкуються за принципом крис-крос. Ознаки, зчеплені з Y-хромосомою, лише по чоловічій лінії й називаються голандричними. Незважаючи на те, що Х- та Y-хромосоми ссавців суттєво відрізняються за набором генів, у дистальних районах короткого та довгого плечей вони мають гомологічні ділянки . псевдоаутосомні регіони.Гени, які знаходяться в цих ділянках (приблизно 10), формально зчеплені зі статтю (містяться в статевих хромосомах), але характер спадкування мають такий самий, як і аутосомні.
Докази реальності еволюційного процесу.
Докази еволюції тісно переплітаються з методами вивчення еволюційного процесу та надані багатьма науками.
Палеонтологічні докази є чи не найпереконливішими документами протікання еволюційного процесу великого масштабу. по-перше, це перехідні форми – форми організмів, що об’єднують ознаки більш стародавніх та більш молодих груп. Пошуки та детальні описи таких форм є важливими методами відновлення філогенезу окремих груп. Яскравий представник – викопна кистепера риба Eusthenopteron, що дозволяє зв’язати риб з наземними хребетними.
Філогенетичний ряд – ряд викопних форм, послідовно зв’язаних одна з одною у процесі еволюції. По кількості знайдених проміжних ланок такий ряд форм може бути більш чи менш детальним, але у всіх випадках це має бути саме ряд форм, близьких не тільки основним, але і частковим деталям будови і безумовно генеалогічно пов’язаних одна з одною у процесі еволюції.
Філогенетичні ряди конструюються на основі аналіза окремих розрізнених знахідок, що відносяться до різних територій. При цьому завжди залишається ймовірним, що деякі форми, які жили у минулому, випадають через відсутність достатньо показного матеріалу. Але при певних сприятливих умовах зберігаються у викопному стані у одному і тому ж місці усі вимерлі форми групи. При пошаровому аналізі таких відкладів можна отримати істинну послідовність виникнення та зміни форм у еволюції. При аналізі послідовності викопних форм виникає можливість визначити реальну швидкість протікання еволюційного процесу.
Географічне розподілення тварин та рослин по Землі може бути пояснене лише з еволюційних позицій. Чим довше та більше ізольовані у процесі еволюції були окремі ділянки біосфери, тем глибше відміни у їх тваринному та рослинному світі. Ці відміни виникають на основі розвитку комплексу предкових видів в умовах постійної географічної ізоляції.
Переконливими еволюційними доказами слугують факти так званого переривчастого розповсюдження організмів. У історії нашої планети звичайними були випадки, коли організми, яки заселяли великі території, зникали з великої частини попереднього ареалу не пристосувавшись до темпів зміни середовища. Такі види зберігалися лише у деяких обмежених ділянках, що мають умови, близькі до попередніх.
Про флору та фауну свідчать і реліктові форми – окремі види або невеликі групи видів з комплексом ознак, характерних для давно вимерлих груп минулих епох. Серед тварин однією з найбільш яскравих реліктових форм є гатерія – єдиний представник цілого підкласу рептилій. У ній відображені риси рептилій, що жили на Землі десятки мільйонів років тому – у мезозої.
Використання морфологічних (порівняльно-анатомічних, гістологічних та ін..) доказів та методів вивчення еволюції засноване на простому принципі: глибока внутрішня подібність організмів може показати спорідненість форм, що порівнюють. Крім загального співставлення будови організмів різних груп існує декілька спеціальних морфологічних підходів, що дозволяють використовувати морфологічні дані для доказу еволюції та досліджувати шляхи еволюційного процесу. Це дослідження гомології, рудиментарних органів та атавізмів, дослідження порівняльно-анатомічних рядів.
Ембріологія дає конкретні докази протікання еволюційного процесу. До них відносяться: закон Бера – явище зародкової подібності – чим більш ранні стадії індивідуального розвитку досліджуються, тем більше подібності виявляється між різними організмами; та принцип рекапітуляції: у процесі еволюції ніби повторюються (рекапітулюють) багато рис будови предкових форм.
За допомогою систематики створюється філогенетична система організмів. Встановлення систематичного положення тієї чи іншої форми відносно інших форм дає змогу вирішити еволюційні проблеми встановлення генеалогії, шляхів еволюційного розвитку порівняних груп. Незважаючи на те, що між великими природніми групами тварин, рослин та мікроорганізмів, як правило, існують великі розриви, викликані вимиранням проміжних форм, у ряді випадків ми виявляємо перехідні форми. Існування форм, що поєднують у своїй будові ознаки різних типів організації та тому займають проміжне систематичне положення, визначається загальною спорідненістю організмів. Існування проміжних форм у сучасному органічному світі – доказ єдності організації крупних стовбурів дерева життя та єдності їх походження.
Генетика – центральна біологічна дисципліна, що дозволяє відкрити особливості протікання процесу еволюції, виявити його рушійні сили та матеріал. Особливе значення для еволюційного вчення дає розвиток популяційної генетики, що дозволяє відкрити протікаючи у елементарних еволюційних одиницях – популяціях – процеси, які призводять до незворотніх змін частот різних генів – елементарним еволюційним подіям.
Корисними також є дані біохімії (визначення «кровної спорідненості» різних груп) та фізіології (розвиток тих чи інших функцій у процесі філогенезу, порівняння особливостей функціонування структур різних гілок дерева життя).
БІЛЕТ 29