

ЭКЗАМЕН ГИСТА (pdf.io)
.pdfДаша: нервные ткани и НС и билеты 3-7 эмбриология Алина:пищеварительная и эндокринная
Илюша: орг чувств, ссс, дыхательная, кожа и произв ЯрШок: мышечные ткани и эпителий
Список сокращений:
грЭПС - гранулярная эндоплазматическая сеть агр ЭПС - агранулярная эндоплазматическая сеть КГ - комплекс Гольджи ГМК - гладкомышечная клетка АХ - ацетилхолин СПМ - спинной мозг ГМ - головной мозг т/о - только С-д - сперматозоид
ЦТБ - цитотрофобласт
ЭКЗАМЕНАЦИОННЫЕ ВОПРОСЫ (ЛФ)
1
ЭМБРИОЛОГИЯ
1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурно-генетическая характеристика.
Эмбриогенез – часть онтогенеза человека, включающая образование и формирование организма на стадии плода.
Стадии эмбриогенеза: -Оплодотворение и образование зиготы -Дробление и образование бластулы
-Гаструляцию (образование зародышевых листков и комплекса осевых органов) -Гистогенез и органогенез зародышевых и внезародышевых органов) -Системогенез
Прогенез – период развития и созревания половых клеток. Результат – получение гаплоидных клеток, способных к оплодотворению и развитию новых организмов
Сперматозоид состоит из головки и хвоста В головке различают:
*плазмолемму: есть специальные белки для хемотаксиса и для связывания овоцитом II порядка
*акросому: мембранный мешочек с литическими ферментами (гиалуронидаза, коллагеназа и другие)
*ядро: гаплоидное В хвосте выделяют отделы:
//шейка: проксимальная и дистальная (от неё начинается аксонема, которая проходит через все отделы хвоста посередине) центриоли
//промежуточный: цитоскелетные фибриллы (только продольные) + куча митохондрий в виде спирали
//главный: фибриллы в продольном и циркулярном направлениях
//концевой: только плазмолемма
Яйцеклетка (по Кузнецову, некорректный термин, мол у человека нет стадии «яйцеклетка», а то, что мы называем яйцеклеткой, на самом деле называется овоцит I порядка после большого роста или овоцит II порядка после овуляции)
По классификации является вторично олиголецитальной и изолецитальной (небольшое количество желточных гранул равномерно распределено во всём объёме цитоплазмы). Желточные гранулы включат фосфо- и липопротеины. Под плазмолеммой лежат кортикальные гранулы с литическими ферментами, которые нужны для кортикальной реакции
Оболочки яйцеклетки:
=блестящая – слой гликозаминогликанов и гликопротеинов, которые синтезируют фолликулярные клетки и овоцит (у фолликулярных клеток есть отростки, которые проходят в прозрачную оболочку, поэтому её называют лучистым венцом)
=слои фолликулярных клеток: на поздних стадиях развития – зернистая (ближе к овоциту) и пристеночная
=тека
2
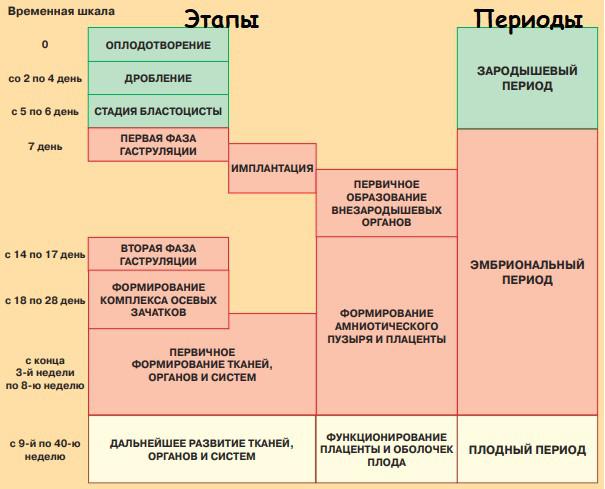
Сперматозоиды истинно гаплоидные, то есть после сперматогенеза каждая клетка nc, а вот овоциты II порядка (которые выходят после овуляции в маточную трубу) по сути не являются гаплоидными: у них всё ещё n2c, а nc они станут только после оплодотворения (если станут)
2. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Сперматогенез и овогенез: стадии и возрастная характеристика.
Стадии сперматогенеза:
() Стадия гоноцитов – клетки, ждущие в стенке желточного мешка своего часа, организм ещё не сделал им домик (семенники) из первичной почки (мезонефроса) [2 месяц эмбрионального развития]
() Пресперматогенез – гоноциты заходят в семенник (они прошли по кровеносным сосудам) и по совету эпителия, находящегося в половых шнурах (на первичной почке), решают стать мужскими половыми клетками. Теперь они - пресперматогонии [3 месяц эмбрионального развития]
() Собственно сперматогенез – начинается репродуктивный возраст, клетки проходят необратимую дифференцировку, становясь при этом сперматозоидами [10-12 лет]
Этапы собственно сперматогенеза:
А) митоз спрематогоний (длительность – 2 недели) Б) мейоз сперматоцитов (длительность – 1 месяц)
В) морфологическое созревание = «формирование» (длительность – 1 месяц) Г) внеэпителиальное созревание в придатке яичка (длительность – 1-3 недели)
Общая продолжительность сперматогенеза – 75 суток (Почему по этапам число выходит больше? Я не знаю)
P.s. Кузнецов разрушил старую этапность:
3
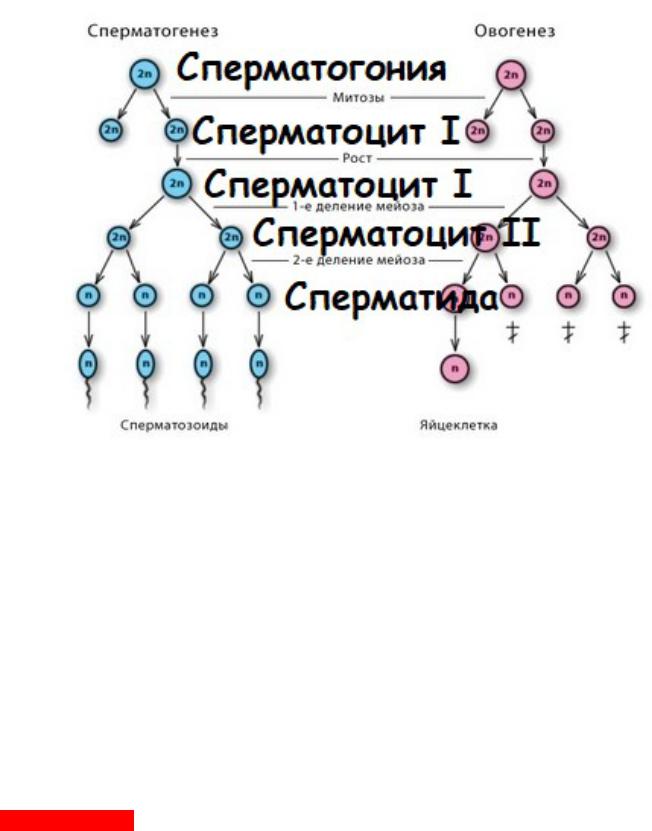
Многие авторы вместо второго из вышеперечисленных этапов (периодов, фаз, стадий) указывают два следующих: период роста и период созревания. При этом под периодом роста понимают длительную профазу 1-го деления мейоза, а под периодом созревания — быстро совершающиеся все прочие фазы обоих делений мейоза. Такая терминология вряд ли удачна: она маскирует важнейшие в биологическом отношении события профазы мейоза предельно упрощенным понятием «период роста»
На всякий случай прикрепляю классику:
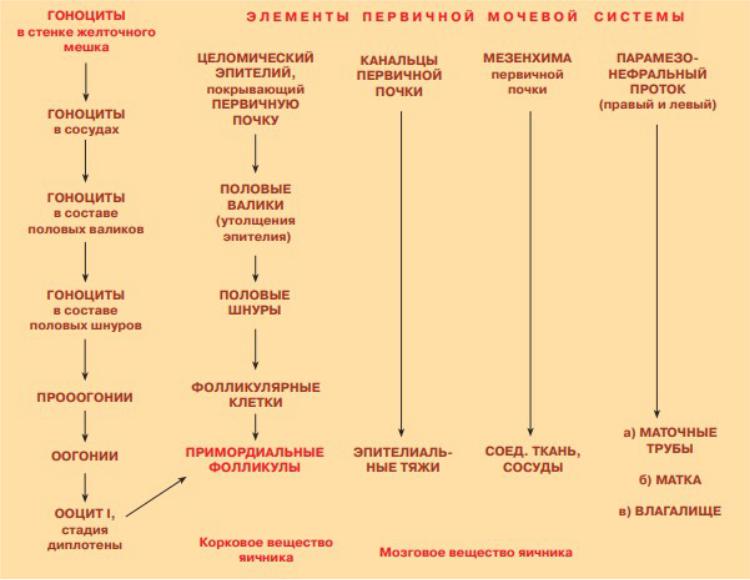
Стадии овогенеза такие же, как и у сперматогенеза:
#Стадия гоноцитов
#Преовогенез
#Собственно овогенез Этапы овогенеза:
^ Митоз – 3-4 месяц эмбрионального развития – образовались овогонии
^Начало профазы мейоза (малый рост) – 3-7 месяц эмбрионального развития – образовались овоциты I, которые доходят до состояния 2n4c и останавливаются
^Покой – от нескольких месяцев (примордиальные фолликулы могут начинать зреть уже у маленьких детей, но они не вызревают) до 50 лет
^Большой рост – всё ещё профаза, но касающаяся в большей степени органоидов - половозрелый возраст
^Постпрофазные деления мейоза – оба мейотических деления – половозрелый возраст (причём овуляция происходит с овоцитом II, который уже начал 2 деление, но остановился в метафазе)
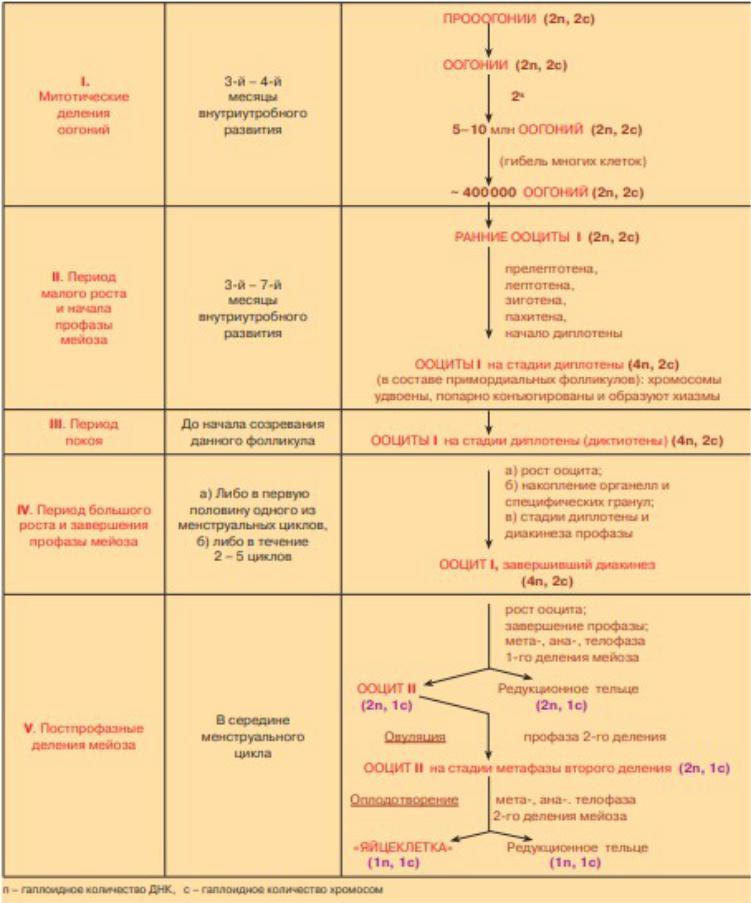
ВНИМАНИЕ!!! Все таблицы Кузнецова, приведённые ниже, используют другую номенклатуру «n» и «c», то есть они переставлены местами по отношению к привычным нам обозначениям
4
5
3. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Понятие оплодотворения. Биомеханизм оплодотворения у человека. Понятие зиготы. Понятие дробления зародыша. Представление о критических периодах развития.
Транспорт овоцита а) из яичника в маточную трубу
6
-перед овуляцией увеличивается тонус гладких миоцитов маточных труб - воронка маточной трубы приближается к поверхности яичника
-увеличивается приток крови к фимбриям, развивается отек их стромы, их тонус увеличивается и они контактируют с яичником
-на эпителиоцитах фимбрий также увеличивается количество ресничек, их биение создает ток жидкости, благодаря которому овоцита 2 порядка переходит с поверхности яичника в маточную трубу
По маточной трубе
-реснички эпителиоцитов маточной трубы тоже начинают биться, мышечная оболочка трубы ритмически сокращается - это происходит под воздействием прогестерона желтого тела * кроме того, что это биение ресничек транспортируют овоцит (они здесь играют не главную роль, в
основном перемещение овоцита происходит из-за сокращений мышечной оболочки), оно еще и указывает сперматозоиду, в какой конкретно яйцевод ему плыть
-секреторные эпителиоциты вырабатывают в-ва, к-ые нужны для питания овоцита
Транспорт сперматозоидов перемещение начинается с момента образования сперматозоида в извитом семенном канальце
! перемещение сперматозоида включает в себя не просто доставку сперматозоида к овоциту, но еще и постоянное дозревание и формирование в каждом отделе семявыносящих путей
извитые семенные канальцы перитубулярные клетки ритмически сокращаются и создают ток жидкости внутри канальцев
прямые семенные канальцы - биение ресничек эпителиоцитов
сеть яичка - биение ресничек эпителиоцитов
выносящие канальцы
-биение ресничек эпителиоцитов
-сокращение их мышечной оболочки
проток придатка
-сперматозоиды попадают сюда и в хвосте придатка остаются здесь 10-15 дней
-эпителий придатка выделяет секрет, к-ый воздействует на мембрану сперматозоидов, модифицирует ее ! именно благодаря этой модификации в протоке придатка сперматозоиды могут активно перемещаться, НО в самом протоке они еще не активны, потому что а) здесь среда слабокислая (в слабокислой среде они очень медленно передвигаются)
б) эпителиоциты вместе с секретом синтезируют иммобилин
эякуляция
-резкое сокращение мышечной оболочки в области хвоста придатка
-сперматозоиды покидают проток придатка
-попадают в семявыносящий проток
-мышечная оболочка семявыносящего протока сокращается
-они перемещаются в уретру
уретра - сперматозоиды смешиваются с продуктами секреции добавочных желез и образуется семенная
жидкость - сперма функции секретов:
а. секрет семенных пузырьков имеет слабощелочную среду = придает подвижность, защищает
7
сперматозоиды в кислой среде уретры и влагалища, содержит фруктозу - важное питательное вещество для с-дов
б. секрет простаты - в нем есть ИГ, гормоны, трансглутаминаза (разжижает эякулят) и т.д. - это все подавляет иммунный ответ женской половой системы на с-ды
Оплодотворение - процесс слияния яйцеклетки и сперматозоида, в результате чего возникает зигота, содержащая диплоидный набор хромосом
фазы:
-фаза дистантного взаимодействия
-фаза контактного взаимодействия
-фаза синкариона (проникновение с-да в овоцит)
-подготовка зиготы к дроблению
фаза дистантного взаимодействия основная задача - обеспечение встречи гамет
впериод овуляции в женском организме существуют механизмы, повышающие и снижающие вероятность встречи гамет - среда слизи в цервикальном канале становится щелочной
- слизь становится жидкой (обычно вязкая)
- гликопротеиновые мицеллы располагаются параллельно оси шеечного канала и создают ходы, по к-ым плывут с-ды (обычно образуют плотную сеть)
вканале шейки матки много с-дов пожирается макрофагами - селекция сперматозоидов
те, к-ые остались подвергаются капацитации (с момента их нахождения в женских половых путях должно пройти 7 часов) - приобретение сперматозоидами оплодотворяющей способности
Одна из гипотез относительно природы капацитации постулирует, что приобретение сперматозоидом оплодотворяющей способности происходит в результате изменения структуры липидов клеточной мембраны сперматозоида.Молекулы альбумина из секрета маточных труб способны связывать холестерин сперматозоида, что изменяет соотношение холестеролфосфолипиды в мембране спермия и мембранный потенциал. Без таких изменений липидного состава слияние наружной мембраны сперматозоида и мембраны акросомы, наблюдающееся при акросомной реакции, не может осуществиться.
Кроме того, при перемещении спермия по маточным трубам с поверхности сперматозоида удаляются углеводные соединения, блокирующие активные центры рецепторных молекул, способных узнавать ZPбелки прозрачной оболочки яйцеклетки.
потом они могут правильно выбрать яйцевод с помощью хемотаксиса - направленное движение сперматозоида по градиенту концентрации веществ, выделяемых яйцеклеткой
-вещества, к-ые выделяют половые клетки называются гамоны
-женские гамоны - гиногамоны увелич. подвижность сперматозоидов и обеспечивают отрицательный реотаксис - движение с-дов против тока жидкости в яйцеводах (противоположное направление слизи тут - ориентир для с-дов)
Фаза контактного взаимодействия Совместное действие трубной жидкости и ферментов (в основном,
пенетразы и гиалуронидазы) некоторого количества сперматозоидов индуцирует разрушение межклеточных контактов фолликулярных клеток
8
лучистого венца. В результате биения жгутиков сперматозоидов яйцеклетка совершает вращательные движения вокруг своей оси. При таком вращении и поступательном движении вдоль трубы складки слизистой удаляют с поверхности овоцита уже не связанные между собой клетки лучистого венца (денудация овоцита).
После удаления лучистого венца обнажается прозрачная оболочка и начинается видоспецифическое взаимодействие половых клеток. Очевидно, что необходимость видоспецифического узнавания гамет сформировалась в процессе эволюции, когда при внешнем оплодотворении в водной среде оказываются половые продукты представителей многих видов. Для осуществления встречи сперматозоидов и яйцеклеток одного вида при столь низкой их концентрации выработались не только механизмы видоспецифического привлечения, но и узнавания гамет с помощью рецепторов.
Фермент галактозилтрансфераза на поверхности сперматозоида (в сперматозоидах, не прошедших капацитации, активные центры этого фермента блокированы) соединяется с концевыми остатками N- ацетилглюкозамина — частью гликопротеина ZP3 прозрачной оболочки овоцита. Видимо, видовую специфичность обеспечивают также биндин и акрозин. Взаимодействие гликопротеинов ZP3 с рецепторами сперматозоида приводит к развитию акросомной реакции. В ходе этой реакции наружная акросомальная мембрана сливается с прилежащей плазмолеммой сперматозоида, что приводит к появлению многочисленных пор, через которые происходит высвобождение акросомальных ферментов. Установлено, что акросомальный экзоцитоз является кальцийзависимым процессом.
Предполагается, что взаимодействие рецепторов гамет приводит к активации вторых посредников — Rabбелков. Они, в свою очередь, активируют белок наружной акросомальной мембраны — синаптотагмин VI, который принадлежит к семейству кальций-связывающих белков. Высвобождение кальция запускает процесс слияния мембран и выход акросомальных ферментов: гиалуронидазы, протеаз
(в т. ч. акрозина — основного фермента, способствующего прохождению сперматозоида через блестящую оболочку), гликозидазы, липазы, нейроминидазы и кислой фосфатазы (способствует прохождению через цитолемму овоцита).
Пик ЛГ, стимулирующий овуляцию, приводит к лютеинизации фолликулярных клеток и синтезу ими прогестерона. При овуляции прогестерон вместе с фолликулярной жидкостью обнаруживается в месте оплодотворения. Выяснилось, что этот гормон оказывает влияние на многие функции сперматозоида: подвижность, капацитацию, способность связываться с рецепторами блестящей оболочки, акросомную реакцию.
Обнаружено два типа рецепторов для прогестерона на поверхности сперматозоида, аналогичных ядерным рецепторам. У сперматозоидов с нарушенной чувствительностью к прогестерону нарушена и оплодотворяющая способность. После выхода ферментов частью наружной оболочки спермия становится мембрана акросомы. В результате ферментативного воздействия на прозрачную оболочку в ней формируется узкий канал, по которому сперматозоид вплотную подходит к цитолемме овоцита. Для слияния мембран гамет вновь необходимо рецепторное объединение между половыми клетками.
В ходе фазы контактного взаимодействия последовательно формируются два типа контактов: сначала между наружной мембраной сперматозоида и прозрачной оболочкой, затем — между оволеммой и бывшей акросомальной мембраной, ставшей частью наружной мембраны спермия после акросомальной реакции. Если первый этап обеспечивает только связывание гамет (в основном, лишь за счет рецепторов, которые относятся к селектиновой группе молекул клеточной адгезии — МКА), то второй — и связывание, и слияние (в основном, за счет кадгеринов, интегринов и молекул ADAMs).
Получается, что с помощью адгезионных молекул устанавливается контакт между гаметами. Этот контакт вызывает, очевидно, полимеризацию актина кортикальной зоны овоцита и увеличение размеров микроворсинок на его поверхности, которые формируют воспринимающий бугорок. В этом участке происходит слияние плазматических мембран овоцита и одного из спермиев. Следует помнить,
что в конце акросомальной реакции переднюю часть головки сперматозоида покрывает внутренняя акросомальная мембрана, и именно она последовательно сливается с микроворсинками овоцита. С этого момента начинается ряд событий, назначение которых — предупреждение полиспермии и создание определенной среды для развития эмбриона. К таким событиям относятся: смена заряда на поверхности яйцеклетки, кортикальная реакция, образование оболочки оплодотворения.
9
В течение одной десятой секунды после прикрепления первого сперматозоида мембрана яйцеклетки деполимеризуется и мембранный потенциал изменяется от –70 мВ до 0 или до +20 мВ. Причина этого явления не ясна. Предположительно, существует акросомный белок, открывающий Nа-
каналы в мембране овоцита. При снижении мембранного потенциала овоцит утрачивает способность взаимодействовать с мембраной другого сперматозоида. Кроме этого, т. н. быстрого блока полиспермии, который поддерживается около минуты, существует и медленный блок, который реализуется посредством кортикальной реакции. Механизм кортикальной реакции сходен с механизмом акросомальной реакции. В результате увеличения концентрации свободных ионов Са2+ в яйцеклетке, мембраны кортикальных гранул сливаются с оволеммой, тем самым вызывая экзоцитоз их содержимого. Экзоцитоз кортикальных гранул начинается вокруг места проникновения сперматозоида, откуда волной распространяется по кортикальному слою и завершается на противоположном полюсе яйцеклетки. Высвободившиеся из кортикальных гранулпротеолитические ферменты разрушают белки, которые связывают плазмолемму яйцеклетки с блестящей оболочкой. В ней происходит протеолиз или модификация ZP-рецепторов сперматозоидов, что надежно блокирует полиспермию. Выделенные мукополисахариды создают осмотический градиент, который привлекает воду в пространство между плазмолеммой и оболочкой. Формируется жидкое перивителлиновое пространство, имеющее характерный ионный состав — необходимая среда для эмбриона. Наконец, выделяется вещество, способствующее уплотнению отделившейся прозрачной оболочки, которая получает название оболочки оплодотворения. Кортикальная реакция является первым этапом активации яйцеклетки, которая заключается в серии биохимических изменений: возрастает потребление кислорода, усиливается углеводный обмен, начинается интенсивный синтез белка с помощью заранее заготовленных, но заблокированных в информосомах мРНК и рРНК. Возрастает обмен фосфата, калия и кальция. Яйцеклетка активизируется. Вместе с активацией метаболизма при контакте со сперматозоидом внутри яйцеклетки происходит перераспределение органелл, включений и морфогенетических детерминант, которое получило название сегрегации цитоплазмы. Эти перемещения приводят к биохимической неоднородности цитоплазмы и играют большую роль при последующем развитии, ведь, попадая в определенные участки зиготы, разные мРНК и белки обеспечивают биохимическую неоднородность этих локусов.
Фаза синкариона Фаза включает набухание и сближение ядер гамет. Она начинается с момента проникновения в овоцит
головки и шейки сперматозоида. Ядро сперматозоида с высококонденсированным хроматином начинает набухать, хромосомные нити раскручиваются — формируется мужской пронуклеус. К этому моменту в овоците заканчивается второе деление мейоза, овоцит становится яйцеклеткой и начинается формирование женского пронуклеуса (ядро набухает, хромосомы из резко конденсированного состояния переходят в диффузное). Наконец пронуклеусы сближаются (синкарион). У человека пронуклеусы не сливаются в единое ядро. В процессе сближения в каждом пронуклеусе происходит удвоение ДНК (хромосомы, находясь в диффузном состоянии, приобретают двухроматидную структуру). Когда пронуклеусы приходят в соприкосновение, их оболочки разрушаются, хромосомы конденсируются, образуют единую материнскую звезду — начинается первое митотическое деление.
Сперматозоид, кроме своей половины хромосом диплоидной зиготы, вносит:
−сигнальный белок дробления. В зиготе обнаружены два принесенные сперматозоидом белка с одинаковой антигенной детерминантой. Антитела к этим белкам блокируют первые дробления зиготы
−проксимальную центриоль, которая играет большую роль в дроблении зиготы. Дупликация центриоли происходит на стадии пронуклеусов. Наследование дефектной центриоли приводит к аномалии дробления и гибели концептуса. При такой патологии неэффективны искусственные методы оплодотворения
−митохондрии — как и любые другие, имеют собственную ДНК. Известны наследуемые с митохондриями заболевания, которые могут иметь как материнское, так и отцовское происхождение.
10