

ЭКЗАМЕН ГИСТА (pdf.io)
.pdf●В процессе сокращения происходит скольжение толстых и тонких миофиламентов относительно друг друга, путем попеременного замыкания и размыкания мостиков между ними (происходит изменение длины)
●Содержат много митохондрий
●Имеют запасы углеводов в виде гранул гликогена и запасы жиров в виде липидных капель
●АТФисточник энергии при сокращении миофибрилл
Происхождение гладкой мышечной ткани
ГМТ сосудов и внутренних органов- мезенхима ГМТ нейрального происхождения (радужка глаза)- клетки нейрального зачатка
Миоэпителиальные клетки экзокриных желез- эктодерма
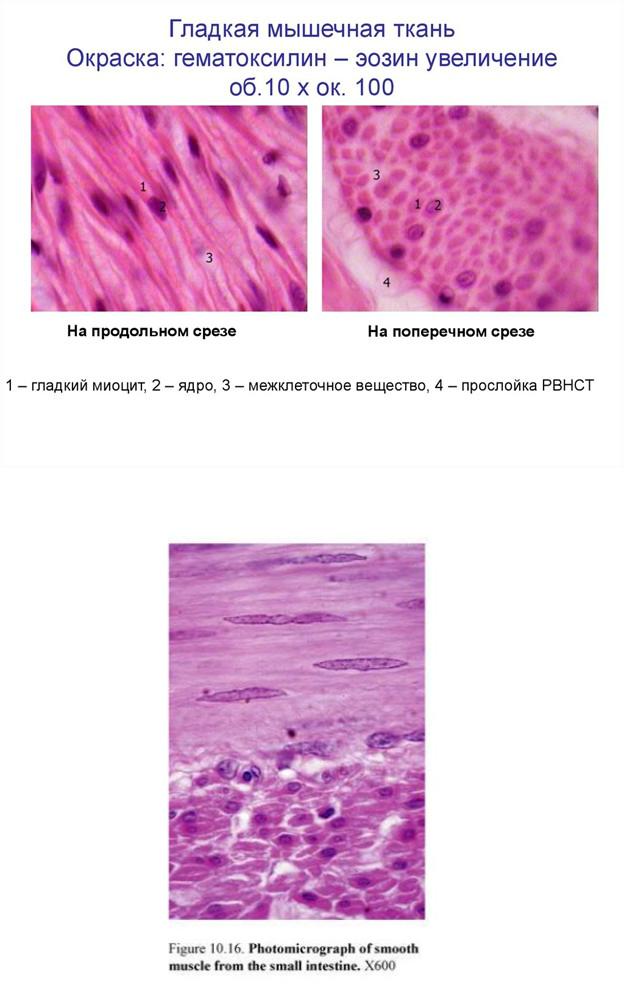
Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и сосудов является миоцит. Представляет собой чаще всего веретенообразную клетку (длиной 20—500 мкм, диаметром 5—8 мкм), покрытую снаружи базальной пластинкой, но встречаются и отростчатые миоциты. В центре располагается вытянутое ядро, по полюсам которого локализуются общие органеллы: зернистая эндоплазматическая сеть, пластинчатый комплекс, митохондрии, цитоцентр. В цитоплазме содержатся толстые (17 нм) миозиновые и тонкие (7 нм) актиновые миофиламенты, которые располагаются в основном параллельно друг другу вдоль оси миоцита и не образуют А и I диски, чем и объясняется отсутствие поперечной исчерченности миоцитов. В цитоплазме миоцитов и на внутренней поверхности плазмолеммы встречаются многочисленные плотные тельца, к которым прикрепляются актиновые, миозиновые, а также промежуточные филаменты. Плазмолемма образует небольшие углубления — кавеолы, которые рассматриваются как аналоги Т-канальцев. Под плазмолеммой локализуются многочисленные везикулы, которые вместе с тонкими канальцами цитоплазмы являются элементами саркоплазматической сети.
Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует выпячивания- кавеолы, в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков. Это влечет за собой взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и « миофибриллы» распадаются. Сокращение прекращается. Гладкая МТ иннервируется вегетативной нервной системой, т.е. не подчиняется воле человека. Сокращение ГМТ медленное - тоническое, зато ГМТ малоутомляема.
Регенерация ГМТ:
1.Митоз миоцитов после дедифференцировки: миоциты утрачивают сократительные белки, исчезают митохондрии и превращаются в миобласты. Миобласты начинают размножаться, а потом вновь дифференцируются в зрелые леомиоциты.
2.Возможно образование новых ГМ-клеток из малодифференцированных стволовых клеток фибробластического дифферона рыхлой с.д. Физиологическая регенерация
51
гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается. Не исключена, однако, и пролиферация клеток
52
53
2. Морфофункциональная характеристика и классификация мышечных тканей. Поперечнополосатая скелетная мышечная ткань: источник развития, строение, иннервация. Структурные основы сокращения мышечного волокна. Типы мышечных волокон. Регенерация.
Скелетная мышечная ткань
+
Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты.
Строение
Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной.
Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями
Z). Он включает анизотропный диск и две половины изотропных дисков(анизотропные A-
диски и изотропные I-диски). - по одной половине с каждой стороны. Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов.
Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что в результате взаимодействия актина и миозина в присутствии кальция и АТФ тонкие нити вдвигаются в промежутки между толстыми без изменения их длины. При этом ширина анизотропных дисков не меняется, а ширина изотропных дисков и полос Н - уменьшается. Строгая пространственная упорядоченность взаимодействия множества толстых и тонких миофиламентов в саркомере определяется наличием сложно организованного
поддерживающего аппарата, к которому, в частности, относятся телофрагма и мезофрагма. Кальций выделяется из саркоплазматической сети, элементы которой
оплетают каждую миофибриллу, после поступления сигнала с сарколеммы по Т- трубочкам (совокупность этих элементов описывается как саркотубулярная система).
54

Миосателлитоциты - это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Скелетная мышечная ткань
●Образует скелетные мышцы
●Составляет 20-25% от общей массы тела
●Иннервируется ЦНС
●Основной элементмышечные волокна (миосимпласт и миосателиоциты), которые окружены базальной мембраной
●Диаметр 50-70 мкм
●Ядра по периферии, под плазмолеммой
●Поперечная исчерченностьчередование светлых и темных полосок
типы мышечных волокон в скелетной мышце - разновидности мышечных волокон с определенными структурными, биохимическими и функцио нальными различиями. Типирование мышечных волокон производится на препаратах при постановке гистохимических реакций выявления ферментов - например, АТФазы, лактатдегидрогеназы (ЛДГ), сукцинатдегидрогеназы (СДГ) (рис. 91) и др. В обобщенном виде можно условно выделить три основных типа мышечных волокон, между которыми существуют переходные варианты.
55

Тип I (красные) - медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами,
высокой активностью окислительных ферментов (например, СДГ), низкой активностью гликолитических ферментов и миозиновой АТФазы, преобладанием аэробных процессов, высоким содержанием пигмента миоглобина (определяющим их красный цвет), крупных митохондрий и липидных включений, богатым кровоснабжением. Численно преобладают в мышцах, выполняющих длительные тонические нагрузки.
Тип IIВ (белые) - быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гликолитических ферментов (например, ЛДГ) и АТФазы, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, относительно низким содержанием мелких митохондрий, липидов и миоглобина (определяющим их светлый цвет), значительным количеством гликогена, сравнительно слабым кровоснабжением. Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип IIА (промежуточные) - быстрые, устойчивые к утомлению, с большой силой, оксилительно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и IIB.
3. Морфофункциональная характеристика и классификация мышечных тканей. Поперечнополосатая сердечная мышечная ткань: источник развития, структурнофункциональная характеристика. Регенерация.
Собственно сердечная, мышечная ткань по своим свойствам занимает промежуточное положение между гладкими мышцами внутренних органов и поперечнополосатыми (скелетными). Она сокращается быстрее гладких, но медленнее поперечнополосатых мышц, работает ритмично и мало утомляется.
56
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов "конец в конец", "конец в бок" формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Строение сердечной мышечной ткани.
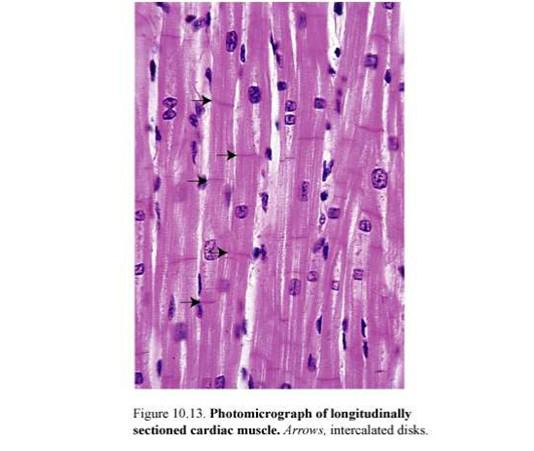
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
В связи с этим в ее строении имеется ряд своеобразных черт .Состоит эта ткань из отдельных мышечных клеток (миоцитов), почти прямоугольной формы, расположенных
столбиком друг за другом. В целом получается структура, напоминающая
поперечнополосатое волокно, разделенное на отрезки поперечными перегородками - вставочные диски,. Рядом лежащие волокна соединены анастомозами, что позволяет им
сокращаться одновременно.
Структурно-функциональной единицей является клетка —кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
·типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
·атипичные кардиомиоциты составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности. Синусные Кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние-проводящим
57
Гистогенез Источники развития поперечно-полосатой мышечной ткани-симметричные участки висцерального листка спланхнотома в шейной части зародыша-
миоэпикардиальные пластинки.Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 5 видов кардиомиоцитоврабочие, синусные переходные, проводящие, а также секреторные.
Регенерация сердечной мышечной ткани. В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности. В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
Сердечная мышечная ткань
●Состоит их двух или одноядерных кардиомиоцитов
●Имеет поперечную исчерченность цитоплазмы
●Автоматияспособность ритмично сокращаться и расслабляться под действием возбуждения
●Выделяют 5 кардиомиоцитоврабочие, синусные, переходные, проводящие, секреторные
58
НЕРВНАЯ ТКАНЬ И НЕРВНАЯ СИСТЕМА
1. Морфофункциональная характеристика нервной ткани. Источники развития. Нейроны (нейроциты): функции, строение, морфологическая и функциональная классификации.
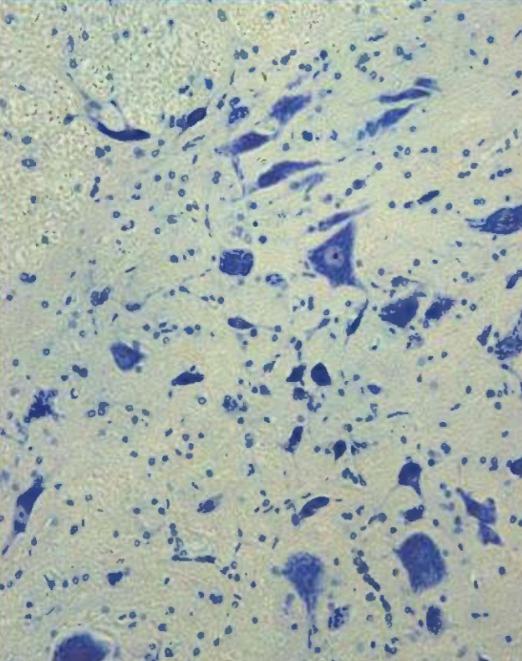
Нервная ткань состоит из а. нейронов - это стабильная популяция
-нет пролиферации б. клеток глии
-растущая популяция
-это значит, что они способны к пролиферации и, если есть сигнал, то клетки глии пролиферируют и, например, замещают дефекты нервной ткани = образуют глиальные рубцы
Итого, мы видим в нервной ткани ядра нейронов, ядра клеток глии, а что между?
Между межклеточное (аморфное) в-во - коллоидный раствор метаболитов -, волокон там никаких нет ! свойства нервной ткани - возбудимость и проводимость
59
источники развития нервной ткани
-нейроэктодерма образует нервную пластинку на 18 сутки
-на 20 сутки формируется нервный желобок, по его бокам находятся нервные валики
-на 21 - н.желобок начинает смыкаться в нервную трубку
-окончательное замыкание происходит на 27-28 сутки, при этом формируются нервная трубка и нервный гребень (ганглиозная пластинка), у нервной трубки остается 2 нейропора. Через них циркулирует амниотическая жидкость
-в головной части рядом с нервной трубкой формируются утолщения эктодермы - нейральные плакоды
И вот эти три образования - источники развития нервной ткани!!
-нервная трубка
-ганглиозная пластинка
-нейральные плакоды
Из нервной трубки – нейроны и глия ЦНС (СПМ и ГМ)
Из нервного гребня - нейроны и глия ПНС (вегетативные и чувствительные ганглии), сенсорные клетки (напр, сенсорные клетки сетчатки глаза, клетки Меркеля), мягкая и паутинная оболочки мозга Материал эктодермальных плакод – ганглии 5, 7, 9, 10 ч/м нервов
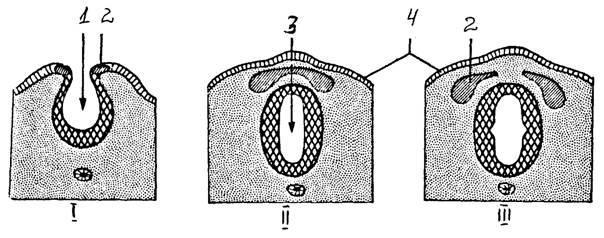
I - образование нервной бороздки, ее погружение |
1 |
- нервный желобок |
II - образование нервной трубки, нервного гребня |
2 |
- нервный гребень |
III - миграция клеток нервного гребня |
3 |
- нервная трубка |
развитие нервной трубки |
4 |
- эктодерма |
|
|
-сначала нервная трубка образует эпителиоподобную структуру
-во внутреннем слое, рядом с невроцелем находятся стволовые клетки - медуллобласты
-они дают начало нейробластам и глиобластам - это 2 основных дифферона нервной ткани
-так вот в этом внутреннем слое (эпендимный слой) клетки размножаются и мигрируют в следующий слой
-плащевой
-в плащевом слое образуются нейробласты и глиобласты
-дифференцировка продолжается, формируются нейроциты и глиоциты
-в последнем слое - краевая вуаль - находятся отростки этих нервных клеток
60