

Нормальная физиология / КР4 кровь + эндокринка
.pdf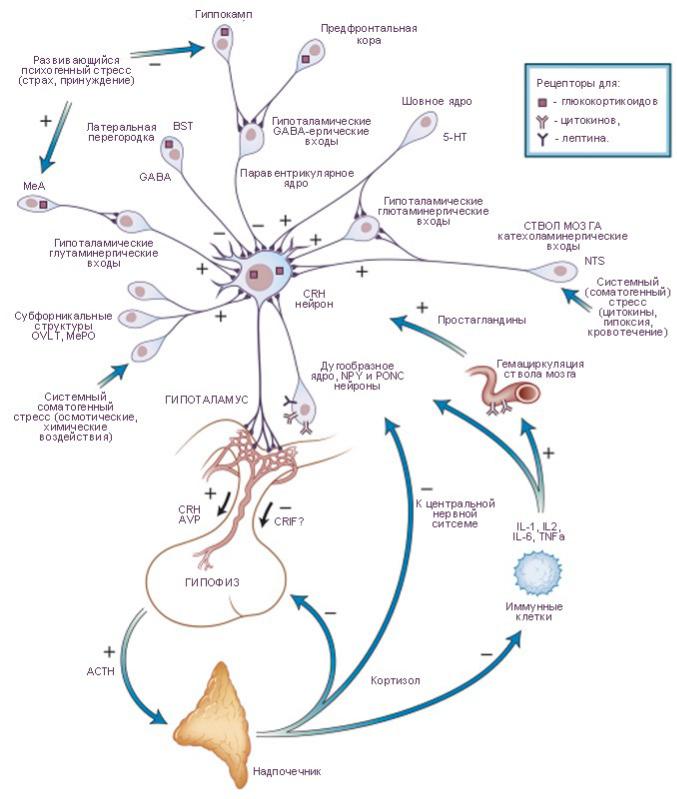
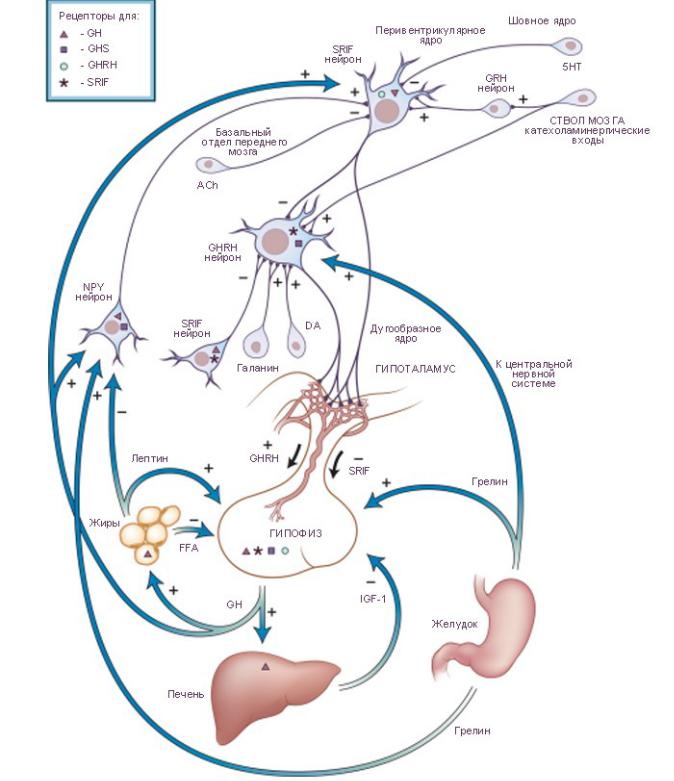
Секреция гормона роста (GH, growth hormone) гипофизом активируется соматолиберином (GH-releasing hormone, GHRH) и тормозится соматостатином (somatostatin, SRIF). На уровне гипофиза существует управление секрецией гормона роста посредством инсулиноподобного фактора роста-1 (insulin-like growth factor I, IGF-I) и посредством свободных жирных кислот (free fatty acids, FFA). Гормон роста также активирует нейроны (SRIF neurons) перивентрикулярного
ядра гипоталамуса, секретирующих соматостатин. Таким образом реализуется механизм короткой отрицательной обратной связи в управлении секрецией гормона роста. Аксоны нейронов, секретирующих соматостатин, заканчиваются синапсами на нейронах (GHRH neurons) дугообразного ядра, вырабатывающих соматолиберин и посылающих свои аксоны к срединному возвышению гипоталамуса. Нейроны дугообразного ядра также непосредственно модулируют секрецию гормона роста
по интегрированным сигналам периферического гормона роста, лептина и грелина. Эти нейроны дугообразного ядра, вырабатывают нейропептид-Y (neuropeptide Y, NPY neurons) и реализуют его к перивентрикулярным нейронам (periventricular SRIF neurons), секретирующим соматолиберин. Грелин секретируется в желудке и является
гипофизарным естественным лигандом для биохимических рецепторов, участвующих в активации секреции гормона роста, как на гипоталамическом, так и на гипофизарном уровне. Установлено, что выведение соматолиберина активируется галанином, γ-аминомасляной кислотой (γ-aminobutyric acid, GABA), и α-2-адренергическими и дофаминергическими (DA, dopamine) сигналами, но тормозится соматостатином. Секреция соматостатина тормозится ацетилхолином (ACh, acetylcholine, мускариновые рецепторы) и 5-гидрокситриптамином (серотонин, 5-HT, 5-hydroxytryptamine, рецепторы типа-1D). Секреция соматостатина активируется α-2-адренергическими сигналами и кортиколиберином.
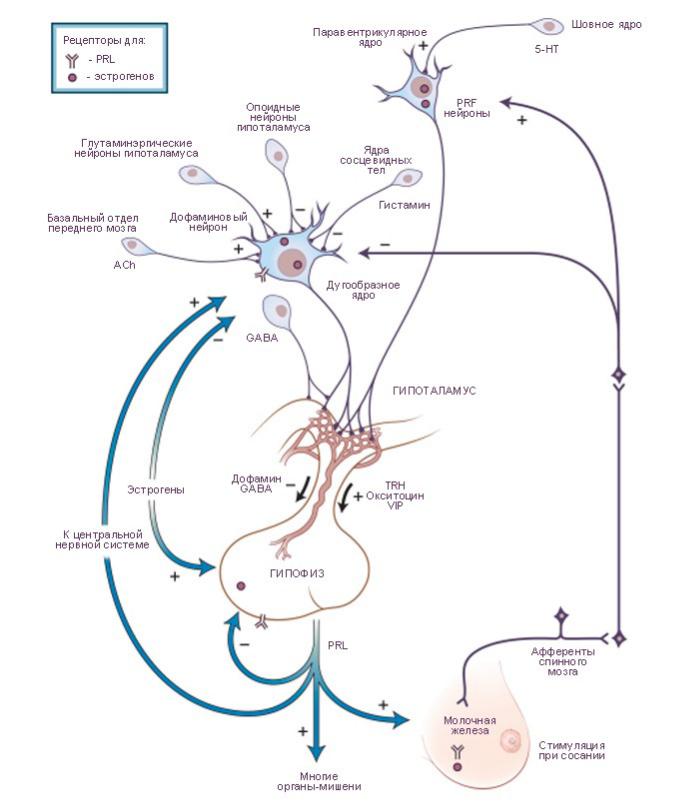
Преимущественное тормозное влияние гипоталамуса на секрецию пролактина запускается дофамином, секретируемого совокупностью
дофаминергических нейронов туберогипофизальной области. Дофаминергические нейроны стимулируются ацетилхолином (acetylcholine, ACh) и глютаматом, но тормозятся гистамином и опоидными пептидами. Немедленное выведение пролактина при сосании или стрессе запускается одним или несколькими пролактолиберинами (prolactin-releasing factors, PRFs). Существует несколько таких пролактолиберинов. Среди них: тиролиберин (thyrotropinreleasing hormone, TRH), тонкокишечный вазоактивный пептид (vasoactive intestinal polypeptide, VIP) и окситоцин (oxytocin).
Нейроны, секретирующие пролактолиберин, активируются серотонином (5-HT). Секреция пролактина также управляется эстрогенами (ультракороткая петля обратной связи). Эстрогены кроме того влияют на секрецию гонадотропина и
подавляют выведение люлиберина (luteinizing hormone-releasing hormone, LHRH). В свою очередь пролактин
по механизму короткой обратной связи влияет на гипоталамический синтез и секрецию дофамина. GABA, γ-aminobutyric acid - гаммааминомасляная кислота.
ПРО КИССПЕПТИН (у человека активен кисспептин 54):
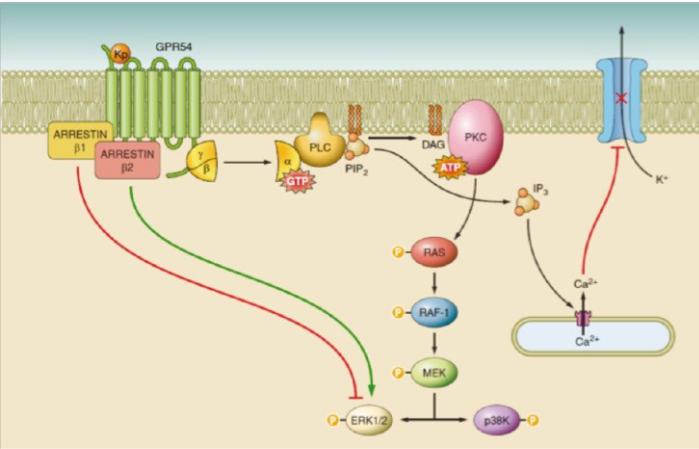
Взаимодействие кисспептина с рецептором сопровождается активацией Gбелков и увеличением уровня внутриклеточного Ca2+ с последующей внутриядерной активацией синтеза ГнРГ и ингибированием пролиферации клеток (поэтому он также является супрессором опухолевого роста)
Взаимодействие кисспептина с рецептором GPR54 приводит к активации Gq-белков и увеличению уровня внутриклеточного Ca2+. Это способствует активации фосфолипазы С (PLC), гидролизу фосфотидилинозитолбисфосфата (PIP2) в инозитол-1,4,5-трифосфат (IP3), стимуляции синтеза диациглицерола (DAG). Вторичные мессенджеры (IP3 и DAG) индуцируют открытие лигандзависимого Са2+-канала, мобилизацию Ca2+ из внутриклеточного депо, активность протеинкиназы С (PKC), фосфорилирование митоген-активируемых протеинкиназ (МАРК) ERK1/2 и р38K, ответственных за усиление внутриядерного синтеза гонадотропин-рилизинг-гормона (ГнРГ) и ингибирование пролиферации клеток.
Примечание: ATP (adenosintriphosphate) ― аденозинтрифосфат; GTP (gamma-glutamyl transferase) ― гамма-глютамилтранспептидаза; MEK ― киназа, активирующая МАРК; Raf ― серин-треониновые киназы; Ras ― семейство генов и белков, которые кодируют G-протеины.
Роль кисспептина в регуляции секреции гонадотропинов
Исследования подтвердили, что кисспептин усиливает секрецию ЛГ и ФСГ как у женщин, так и у мужчин. Стимулирующее воздействие кисспептина на ГГЯО (гипоталамо-гипофизарно-яичниковой оси) было выявлено при различных физиологических состояниях, в различные фазы менструального цикла, а также при беременности и лактации.
Роль кисспептина в половом созревании Среди причин идиопатического гипогонадизма приблизительно 2% составляют инактивирующие
мутации гена кисспептина и его рецептора. В экспериментальных исследованиях показано, что KISS1Rнокаутные мыши не достигают половой зрелости изза отсутствия секреции ЛГ и активности гена Сfos (маркера клеточной пролиферации, дифференцировки и нейрональной активности). В то же время активирующие мутации гена KISS1 и его рецептора связаны со стимуляцией сигнального пути кисспептина и преждевременным половым созреванием.

№ 9. Гормоны аденогипофиза, их физиологическая роль, регуляция секреции
Деятельность гипофиза регулируется гипоталамусом. Действуя совместно, их гормоны регулируют все аспекты роста, развития, обмена веществ и гомеостаза.
Гипофиз —небольшая железа около 1 см в диаметре и массой 0,5-1 г, которая лежит в гипофизарной ямке турецкого седла клиновидной кости. Имеет форму горошины и связан с гипоталамусом при помощи воронки гипоталамуса — infundibulum- и состоит из 2 различающихся частей: передней и задней долей. Передняя доля гипофиза, аденогипофиз, составляет около 75% всей массы железы. У взрослого человека передняя доля гипофиза подразделяется на две части: большую — дистальную (pars distalis) и бугорную часть (pars tuberalis), уходящую вверх и соединяющуюся с воронкой гипоталамуса.
В эмбриогенезе две доли гипофиза развиваются из разных источников: передняя доля — из кармана Ратке, который эмбриологически является инвагинацией фарингеального
эпителия, задняя доля — из нервной ткани, являющейся выростом гипоталамуса.
Передняя доля гипофиза
Передняя доля гипофиза, или аденогипофиз (адено — железа, гипофиз — подрост), вырабатывает 5 пептидных гормонов.
Структура передней доли гипофиза представлена 8 типами клеток, однако секреторная функций присуща только хромафильным. Выделяют 5 типов хромафильных клеток.
Передняя доля гипофиза продуцирует:
•гормон роста (соматотропный гормон, соматотропин) обеспечивает рост, увеличение количества клеток и их дифференцировку, стимулируя образование белков, вырабатывается ацидофильными красными клетками с мелкими гранулами (соматотрофы);
•адренокортикотропный гормон (кортикотропин) управляет секрецией некоторых гормонов коры надпочечников, влияющих на метаболизм глюкозы, жиров и белков, вырабатывают базофильные кортикофы;
•тиреотропный гормон (тиреотропин, тиреоидстимулирующий гормон) контролирует секрецию тироксина и трийодтиронина щитовидной железой, которые регулируют в организме скорость основных химических реакций в клетке, вырабатывают базофильные тиреотрофы;
•гонадотропные гормоны (фолликулостимулирующий и лютеинизирующий) контролируют рост яичников и яичек, их гормональную и репродуктивную активность, вырабатывают базофильные гонадотрофы;
•пролактин обеспечивает развитие молочных желез и продукцию грудного молока, вырабатывают ацидофильные жёлтые клетки с крупными гранулами (лактотрофы).
Регуляция секреции гормонов передней доли гипофиза
2 принципа:
1)Иерархический: посредством РГ и статинов, вырабатываемых мелкоклеточными ядрами гипоталамус
2)Принцип обратной связи:
•положительная обратная связь – при увеличении концентрации гормона периферической эндокринной железы увеличивается синтез и секреция соответствующего тропного гормона и рилизинг-гормона (РГ). Обеспечивает процессы роста, развития (например, работа ФСГ и ЛГ при половом созревании)
•отрицательная обратная связь – при увеличении концентрации гормона периферической ЭЖ снижается синтез и секреция соответствующего ТГ и РГ + может стимулироваться выработка статина. Обеспечивет гомеостаз и гомеокинез + адаптивность систем организма (пример: при

субклиническом гипотиреозе при нормальных уровнях Т3 и Т4 наблюдается повышение уровня ТТГ в анализе крови)
Соматотропин (РЕГУЛЯЦИЯ ЧЕРЕЗ Gq-БЕЛОК)
Соматотропин — это белок (из 191 ак), который образуется в передней доле гипофиза. Продуцируется непрерывно и «вспышками»- повышение секреции происходит во время глубокого сна и на ранних его стадиях.
Регуляция синтеза. Продукция стимулируется соматолиберином (вторичный посредник Са2+) и подавляется соматостатином.
Эффекты соматотропина. Основной функцией является стимуляция секреции белковых гормонов — инсулиноподобных факторов роста (ИФР)- IGF1 и IGF2, или соматомединов. Эти пептиды образуются во многих клетках, преимущественно в печени. Большинство эффектов соматотропина (кроме липолиза) осуществляется посредством ИФР – т.е. минерализция костей, рост и развитие тканей.
Важнейшие эффекты соматотропина:
•стимулирует рост костей, мышц и внутренних органов, а также синтез белков (в т. ч. коллагена), необходимых для роста.
•тормозит глюконеогенез, синтез аминокислот и ограничивает затраты глюкозы посредством торможения ее поглощения жировыми и мышечными клетками
•усиливает липолиз в жировой ткани, который приводит к повышению использования жирных кислот для продукции АТФ клетками тела.
•повышает всасывание Nа+ в почках (через IGF1).
•стимулирует образование кальцитриола, который вызывает поглощение Са2+ в кишечнике и всасывание Са2+ и фосфата в почках. При этом возникают условия для минерализации костей.
•вызывает пролиферацию клеток во многих тканях, например в хрящах (рост костей) и кроветворных органах (эритропоэз).
•Через стимуляцию Т-лимфоцитов и макрофагов поддерживает иммунную систему
Нарушение синтеза соматотропина
Недостаток соматотропина. Недостатокпри гипофизарной недостаточности или др. Даже при нормальной секреции соматотропина его действие может быть недостаточным, если нарушен синтез IGF в печени (при печеночной недостаточности). Следствием недостатка или пониженной эффективности у ребенка является маленький рост. Недостаток соматотропина у взрослого часто остается недиагностированным. Как правило, снижение концентрации соматотропина способствует преобладанию деградации белков и ослаблению работы иммунной системы в старости.
Избыток соматотропина. Избытокпри развитии опухоли соматотропинпродуцирующих клеток. Как следствие избытка соматотропина в период до завершения роста развивается гигантизм. После прекращения роста (завершения роста эпифизов) длина тела остается неизменной. Вместо этого наблюдается акромегалия, т. е. повышенный аппозиционный рост костей. Особенно заметно увеличение подбородка и носа, а также расширение костей челюсти и скул, рук и ступней ног. Значительно увеличиваются размеры внутренних органов, таких как сердце, печень, почки и щитовидная железа, а также язык (макроглоссия).
Пролактин
Синтез. Пролактин — пептидный гормон (из 199 ак). Синтез стимулируется окситоцином (нет своего РГ) и тормозится дофамином, а также пролактостатином. Влияние дофамина на секрецию пролактина преобладает, т. е. при нарушении влияния гипоталамуса наблюдается усиление высвобождения пролактина.
Секреция пролактина зависит и от уровня в крови эстрогенов, глюкокортикоидов и тиреоидных гормонов, изменяющих число рецепторов пролактолиберина на лактотрофах. Повышается секреция при беременности, кормлении грудью, во время сна, физ нагрузки и стресса.
Эффекты. Пролактин стимулирует рост, дифференцировку и функционирование молочной железы, затормаживает выработку гонадотропинов (ЛГ, ФСГ) и действует (преимущественно ингибирует) на иммунную систему. Его секреция повышается при стрессе.
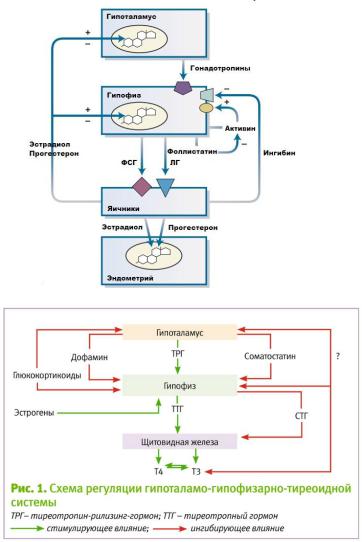
Тиреотропин
Тиреотропин — гликопротеидный гормон аденогипофиза — секретируется непрерывно, с ритмичными колебаниями в течение суток, при этом максимум содержания ввечерние часы.
Секреция тиреотропина стимулируется тиреолиберином, а подавляется соматостатином.
Таким образом, тиреотропин является ведущим звеном гипоталамо-аденогипофи-зарно-тиреоидной регуляторной оси.
•Тормозят секрецию дофамин, глюкокортикоиды и соматотропин, а стимулируют — вазопрессин и антагонисты дофаминовых рецепторов.
•секретируется в повышенных количествах при низкой температуры, другие же воздействия — травма,
боль, наркоз — секрецию гормона подавляют.
Механизм – через Gs-, Gq-белки.
Тиреотропин связывается со специфическим рецептором фолликулярных клеток щитовидной железы и вызывает метаболические реакции с помощью четырех вторичных посредников: цАМФ, инозитол-3-фосфата, диацилглицерола и комплекса Са2+—кальмодулин. Под влиянием тиреотропина в клетках фолликулов щитовидной железы меняются все виды обмена веществ, ускоряется захват иода и осуществляется синтез тиреоглобу-лина и тиреоидных гормонов. Тиреотропин увеличивает секрецию гормонов щитовидной железы активацией гидролиза
тиреоглобулина. Благодаря увеличению синтеза пурина и пиримидина, РНК, белка и фосфолипидов тиреотропин вызывает увеличение массы щитовидной железы.
Внетиреоидное действие тиреотропина проявляется повышением образования гликозаминогликанов в коже и подкожной клетчатке, что ведет к так называемому слизистому отеку, или микседеме.
Клиника
Избыточная секреция тиреотропина приводит к развитию зоба, гиперфункции щитовидной железы с эффектами избытка тиреоидных гормонов (тиреотоксикоз), пучеглазию (экзофтальм), что в совокупности называют «базедова болезнь».
Гонадотропины
Секреция имеет выраженную суточную и месячную цикличность как у мужчин, так и у женщин. Молекулы гонадотропинов секретируются с прикрепленными на конце углеводных цепей гликопротеида сиаловыми кислотами, что защищает их от разрушения в печени.
Регуляция секреции
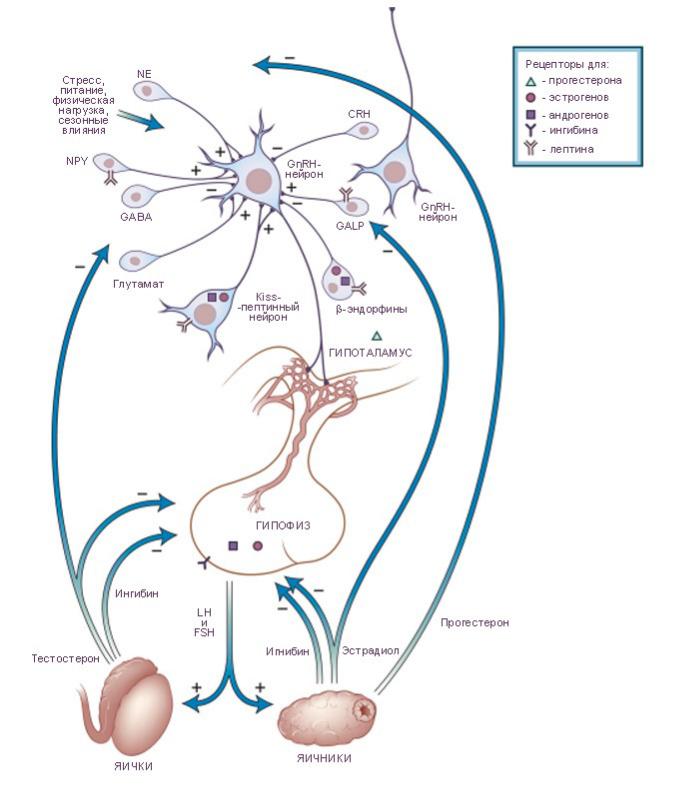
Активация обеспечивается гонадолиберином. Эффект гонадолиберином на секрецию зависит от циклических изменений содержания в крови половых гормонов — эстрогенов, прогестерона и тестостерона (отрицательная обратная связь). ПРИ ЭТОМ СЕКРЕЦИЯ ГОНАДОЛИБЕРИНА ПОДВЕРЖЕНА ВЛИЯНИЮ КИССПЕПТИНА.
Секреция гонадолиберина стимулируется катехоламинами, а подавляется эндогенными опиоидными пептидами, гастрином, холецистокинином и соматостатином. Главный тормозной эффект на продукцию ФСГ оказывает гормон семенников — ингибин. Тормозят секрецию гонадотропинов гормон аденогипофиза пролактин и мелатонин эпифиза; секрецию ЛГ угнетают и глюкокортикоиды.