

Нормальная физиология / КР4 кровь + эндокринка
.pdf
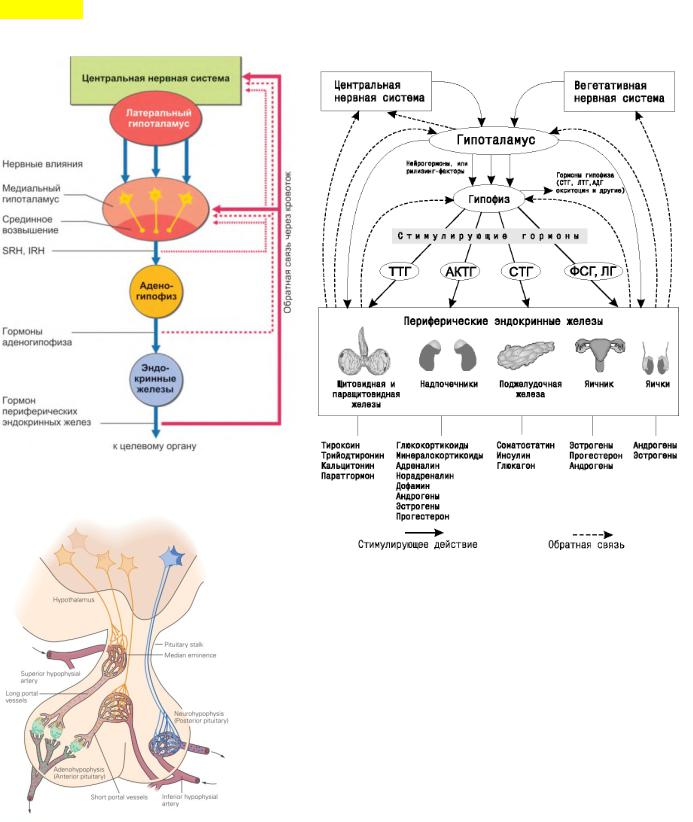
Вопрос 8. Роль гипоталамо-гипофизарной системы в регуляции деятельности периферических ЭЖ.
Гипоталамическая регуляция эндокринной системы.
Гипоталамус управляет периферическими гормонами и контролирует соответствие работы гормонзависимых органов необходимому для данного состояния организма уровню и путем регуляции гормонов аденогипофиза гипоталамическими гормонами (либеринами и статинами). Петли регуляторных контуров гипоталамических, тропных и периферических гормонов устанавливают иерархию.
Регуляторные (гормональные) системы (оси):
1.Гипоталамо-аденогипофизарно- надпочечниковая: кортиколиберин – АКТГ –
глюкокортикоиды;
2.Гипоталамо-аденогипофизарно-гонадная: кисспептин - гонадолиберин – ЛГ и ФСГ
– эстрогены, прогестерон, тестостерон;
3.Гипоталамо-аденогипофизарно-тиреоидная: тиреолиберин – ТТГ – тиреоидные гормоны;
4.Система СТГ: соматолиберин/соматостатин – соматотропин;
5.Система пролактина: окситоцин/дофамин пролактин;
Гипоталамические и тропные гормоны.
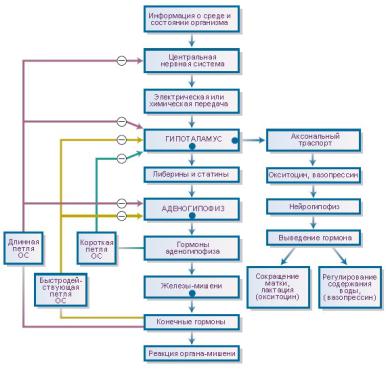
Гипоталамус синтезирует либерины (рилизинг-факторы, или рилизинг-гормоны, РГ) и статины (ингибирующие факторы или гормоны), которые через нервные окончания высвобождаются в воротную систему гипофиза. Сосуды образуют две последовательные капиллярные сети. В первую капиллярную сеть выделяются либерины и статины, вторая сеть капилляров окружает клетки передней доли гипофиза, где синтезируются тропные гормоны.
Рилизинг-гормоны стимулируют секрецию соответствующих тропных гормонов, а ингибирующие (release-inhibiting) гормоны подавляют ее. Рилизинг-гормоны выделяются аксонами нейронов в зоне срединного возвышения и через вены гипоталамо-гипофизарной портальной системы попадают в аденогипофиз.
Мелкоклеточные ядра находятся вдоль стенки 3-го желудочка мозга, содержат нейроны, которые выделяют гормоны в срединное возвышение.
o нейроны, выделяющие гонадотропин-РГ (GnRH), атипичны, т.к. они разбросаны
на протяжении от срединной перегородки до медиобазальных ядер гипоталамуса, при этом их контролируют более вышестоящие нейроны (KISS1-нейроны), выделяющие kisspeptin (кисспептин).
oпулы нейронов, секретирующих остальные РГ, находятся вокруг паравентр. я. (ПВЯ) + есть 2 пула мелкоклеточных нейронов: 1-й выделяет РГ соматотропина, 2-й – дофамин, чтобы ингибировать секрецию пролактина.
oсекреция гормонов идет в систему гипоталамо-гипофизарной воротной системы, которая позволяет им преодолевать ГЭБ и воздействовать на клетки аденогипофиза.
oгонадотропин-РГ, соматотропин-РГ, тиреотропин-РГ – ФОСФОЛИПАЗА С (Gq)
oкортикотропин-РГ – цАМФ (Gs - возбуждение), фосфолипаза С (Gq) + G0 – ионные каналы + цАМФ (Gi – ингибирование).
!Только пролактин контролируется окситоцином и дофаминэргическими нейронами, не имея РФ/ИФ.
Выделение гипоталамических гормонов нейронами в вены портальной системы контролируется концентрацией гормонов периферических эндокринных желез в плазме.
Увеличение концентрации гормонов периферических эндокринных желез в плазме приводит к снижению секреции соответствующих рилизинг-гормонов в медиальном гипоталамусе.
Гипоталамические гормоны и гормоны аденогипофиза также принимают участие в отрицательной обратной связи в этой регуляции.
Петли обратной связи. Параметры обмена веществ, регулируемые периферическими гормонами, оказывают влияние на гипоталамус. Более того, периферические гормоны оказывают ингибирующее влияние на гипоталамус и гипофиз. Наконец, тропный гормон или даже сам рилизинг-гормон может затормаживать собственную секрецию в гипоталамусе.
Адаптация отрицательной нейроэндокринной обратной связи через ЦНС. Гипоталамо-гипофизарная система адаптируется к внутренним и внешним воздействиям на организм через ЦНС. Центральное нервное управление осуществляется через латеральный гипоталамус и прежде всего исходит из преоптической области (region praeoptica), структур лимбической системы (например, гиппокампа и миндалины) и структур среднего мозга. Эти области ЦНС получают и обратную связь от эндокринных желез через концентрацию гормонов в плазме. Нейроны специфически реагируют на эндокринные гормоны и накапливают их внутри клетки.
Регуляция аденогипофиза. Функционирование большинства эндокринных желез управляется гормонами аденогипофиза. Продукция под контролем гормонов, которые вырабатываются нейронами в паравентрикулярной области и медиальном гипоталамусе.
К тропным гормонам передней доли гипофиза относятся:
■Лютеинизирующий гормон (ЛГ) и фолликулостимулирующий гормон (ФСГ). Их выработку стимулирует гонадотропин-рилизинг-гормон (ГнРГ). ЛГ и ФСГ регулируют продукцию половых гормонов — эстрогенов, прогестерона и тестостерона.
■Адренокортикотропный гормон (АКТГ, или кортикотропин). Его синтез стимулируется гормоном кортиколиберином КТ-РГ. АКТГ прежде всего стимулирует секрецию гормонов и рост коры надпочечников.
■Тиреотропный гормон (ТТГ, тиреотропный гормон). Его секреция стимулируется тиреолиберином (ТТГ-РГ). ТТГ стимулирует продукцию гормонов и рост щитовидной железы.
■Соматотропный гормон (СТГ, гормон роста) регулируется соматолиберином (СТГ-РГ) и соматостатином. Основной эффект соматотропина — стимуляция роста, прежде всего через
инсулиноподобные факторы роста (IGF).
Нарушения в работе контролируемых гипоталамусом и гипофизом гормонов возникают при повреждении этих органов. При отсутствии гипоталамического влияния на переднюю долю гипофиза наряду с повышенной секрецией пролактина понижается выброс соматотропина, адено-кортикотропного гормона, меланотронного гормона, тиреотропного гормона и гонадотропинов.
Гипофизарная недостаточность
Гипофиз и гипоталамус могут повреждаться или разрушаться из-за опухолей, воспаления, отложений (например, железа при гемохроматозе), инфекций (например, при туберкулезе), нарушения кровообращения, травм (черепно-мозговые травмы), операций или облучения (например, при терапии опухоли гипофиза). В результате происходит снижение или прекращение секреции нескольких пли всех гипофизарных гормонов.
недостаточность соматотропина – низкорослость у детей.
гонадотропина - отсутствие менструаций, импотенция, снижение либидо и уменьшение оволосения тела.
Т3, Т4 - к анемии, непереносимости холода, брадикардии, остановке и нарушению умственного развития.
глюкокортикоидов - к снижению артериального давления, а также к слабости, гипогликемии, потере веса и высокой температуре
Последствия проявляются медленно при постепенном нарушении функций гипофиза, поэтому долгое время остаются незамеченными. Полное выпадение функций гипофиза летально при отсутствии лечения. В первую очередь проводят заместительную терапию для
периферических гормонов. При этом теряется возможность адаптации уровня синтеза гормона к потребностям организма.
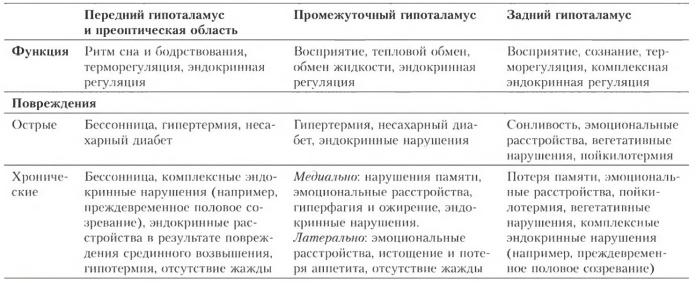
Нарушения функций при повреждениях гипоталамуса у человека
Причины. Нарушения обусловленных гипоталамусом функций у человека вызваны чаще всего неоплазиями (опухолями), травмами и воспалениями. Эти повреждения иногда относительно локализованы, что может приводить к изолированным выпадениям функций в переднем, промежуточном и заднем таламусе.
Симптомы. Дисфункции, наблюдаемые клиницистом у пациента (за исключением несахарного диабета), имеют сложную природу. Они зависят от того, имеет ли повреждение острый (например, из-за травмы) или хронический характер (например, из-за медленно разрастающейся опухоли). Острые небольшие повреждения могут привести к значительным нарушениям функций, в то время как дисфункции из-за медленно растущей опухоли проявляются только тогда, когда повреждения достигают больших размеров.
Гипоталамо-аденогипофизарно-надпочечниковая система
Гипоталамус секретирует кортиколиберин и вазопрессин. По механизму прямых связей они усиливают секрецию кортикотропина гипофизом.
Эффекты кортиколиберина усиливают:
Адреналин и ангиотензин-2;
АХ;
Норадреналин;
Серотонин;
Холецистокинин;
Бомбезин;
Атриопептид;
Ослабляют:
Опиоидные пептиды;
ГАМК.
*кортикотропин – центральное звено этой системы; продукт расщепления крупного 239 АК гликопротеина проопиомеланокортина, образующегося базофильными кортикотрофами. Проопиомеланокортин служит источником кортикотропина, а также липотропина (расщепляясь, дает меланотропин и пептид эндорфин – морфиноподобное действие, роль в антиноцицептивной системе мозга и в модуляции секреции гормонов аденогипофиза).
Секреция кортикотропина происходит пульсирующими вспышками с четкой суточной ритмичностью. Наивысшая концентрация гормона в крови – в утренние часы 6-8, наиболее низкая – с 18 до 2 ночи.
Глюкокортикоиды, циркулирующие в крови (кортизол) в гипоталамусе тормозят секрецию кортиколиберина, в гипофизе – кортикотропиина. То есть обратная связь имеет отрицательную направленность.
Физиологические эффекты кортикотропина: 1. Надпочечниковые:
Основные, заключаются в стимуляции клеток пучковой зоны коры надпочечников, секретирующей глюкокортикоиды (кортизол и кортикостерон). Меньший эффект на продукцию минералкортикоидов и половых стероидов клетками пучковой и клубочковой зон коры надпочечников.
Под влиянием кортикотропина усиливается стероидогенез и синтез белков ткани (перестройка структуры коры, увеличение массы клеток пучковой зоны).
2.Вненадпочечниковые:
Вжировой ткани, действие на другие гормоны и т.п.
Гипоталамо-аденогипофизарно-гонадная система
Гипоталамус секретирует гонадолиберин. Секреция гонадолиберина стимулируется катехоламинами, подавляется – опиоидными пептидами, гастрином, холецичтокинином, соматостатином.
Гонадолиберин активирует синтез фоллитропина и лютропина аденогипофизом. Секреция гонадотропинов имеет четко выраженную суточную и месячную цикличность как у мужчин, так и у женщин. Секрецию гонадотропинов тормозят пролактин и мелатонин, также фоллитропин тормозится ингибином, а лютропин – глюкортикоидами.
Гонадотропины реализуют свое действие через систему аденилатциклазы цАМФ. Влияют на образование и секрецию половых гормонов и на функции половых желез.
Гипоталамо-аденогипофизарно-тиреоидная система
Гипоталамус – тиреолиберин/соматостатин.
Секреция тиреотропина гипофизом стимулируется тиреолиберином, подавляется соматостатином.
Тиреотропин усиливает выработку Т3 и Т4 щитовидной железы.
По механизму отрицательной обратной связи повышение в крови гормонов щитовидной железы ведет к подавлению секреции тиреотропина. Секрецию тиреотропина также
подавляют дофамин, глюкокортикоиды и соматотропин, стимулируют – вазопрессин и антагонисты дофаминовых рецепторов.
Тиреотропин связывается со специфическим рецептором фолликулярных клеток щитовидной железы, вызывает метаболические реакции с помощью 4 вторичных посредников: цАМФ, инозитол-3-фосфата, диацилглицерола и комплекса Са-кальмодулин. Под влиянием тиреотропина в клетках фолликулов щитовидной железы меняются все виды обмена веществ, ускоряется захват йода и осуществляется синтез тиреоглобулина и тиреоидных гормонов. Благодаря увеличению синтеза пурина и пиримидина, РНК, белка и фософлипидов, тиреотропин вызывает увеличение массы щитовидной железы.
Система СТГ
Секреция соматотропина (соматотропного гормона = СТГ) стимулируется соматолиберином (в механизме действия основной посредник – Са), подавляется соматостатином (приводит к снижению концентрации ионов Са в соматотрофах аденогипофиза).
Механизм действия соматотропина на клетки-эффекторы заключается в его связывании с 2 молекулами мембранных рецепторов, активации тирозинкиназы, фосфорилировании и активации цитоплазматических белков.
СТГ участвует в регуляции роста и развитии организма. Основные эффекты СТГ:
1.Усиление липолиза и уменьшение массы жировой ткани;
2.Повышение усвоения АК и синтеза белков, в результате масса тела увеличивается за счет нежировой ткани;
3.Увеличение глюконеогенеза и повышение уровня сахара в крови.
Большинство ростовых эффектов СТГ опосредуется соматомединами – гуморальными факторами печени, почек и костной ткани. Их эффект сходен с эффектом инсулина – инсулиноподобные факторы роста.
Система пролактина
Синтез и секреция аденогипофизом пролактина в основном регулируется гипоталамическим ингибитором дофамином (пролактостатин) и стимуляторами – пролактолиберином и тиреолибеином. Пролактолибериновой активностью также обладают окситоцин, серотонин и вазоинтестинальный пептид. Секреция пролактина зависит от уровня в крови эстрогенов, глюкокортикоидов и тиреоидных гормонов, изменяющих число рецепторов пролактолиберина и тиреолиберина на лактотрофах. Повышается секреция пролактина при беременности, кормлении грудью, во время сна, физической нагрузки, стресса.
Одним из органов-мишеней является молочная железа, где гормон стимулирует развитие секреторной ткани, рост желез, лактацию, оказывая свой эффект после связывания со специфическим рецептором и образования вторичного посредника цАМФ.
В молочных железах пролактин влияет на образование молока, не на его выделение. При этом гормон стимулирует синтез белка – лактальбумина, а также жиров и углеводов молока. Эффект пролактина на лактацию во многом опосредуется образованием в печени лактогенного фактора. Пролактин способствует поддержанию секреторной активности желтого тела в яичниках и образованию прогестерона.
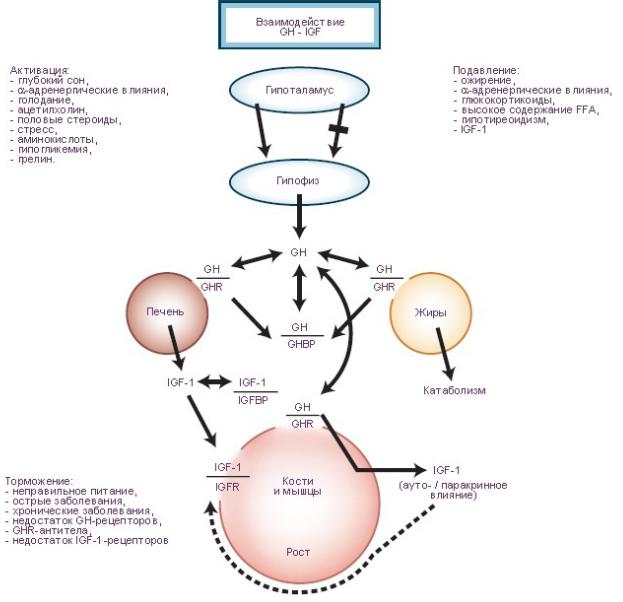
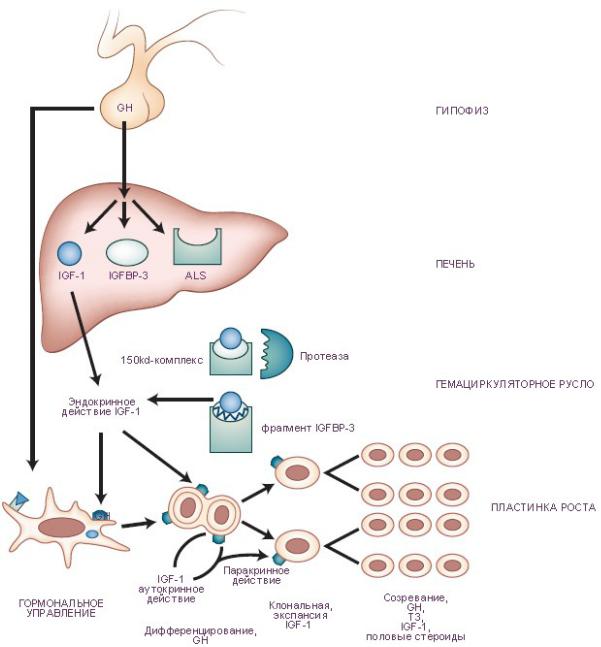
Предполагается существованиt трех возможных механизмов гормонального управления процессами роста.
Всоответствии с первой гипотезой гормон роста (growth hormone, GH) стимулирует выработку инсулиноподобного фактора роста-1 (соматомедин, insulin-like growth factor 1, IGF-1). Повышение концентрации соматомедина в
циркулирующей крови (эндокринный соматомедин) действует на эпифизеальную пластинку (пластинка роста, гиалиновая пластинка хряща в метафизах длинных костей).
Всоответствии со второй гипотезой гормон роста регулирует выработку клетками печени: (а) связанного с белком соматомедина-3 (IGF-binding protein 3, IGFBP-3) и кислотно-лабильных субъединиц (acid-labile subunit, ALS) комплекса белок-
соматомедин (IGFBP complex). Соматомедин связывается с кислотно-лабильными субъединицами комплекса белоксоматомедин. При этом образуется трёхкомпонентные комплексы (150-kd ternary complex). В
последующем протеазы расщепляют эти комплексы на фрагменты, высвобождающие в гемациркуляторное русло связанный
сбелком соматомедин-3 и инсулиноподобноый фактор роста-1, действующие на эпифизеальную пластинку.
Всоответствии со третьей гипотезой гормон роста вызывает дифференцирование и местную выработку инсулиноподобного фактора роста-1 (соматомедин). По паракринному и аутокринному механизму соматомедин стимулирует
деление клеток. T3, triiodothyronine = трииодтиронин.