

Общепринятая модель терморегуляции
Здесь мы изложим несколько более сложную, но значительно более полезную схему. Как и в других ситуациях, когда приходится прибегать к схемам, данную модель следует рассматривать лишь как своего рода каркас, позволяющий выработать подход к анализу терморегуляторных реакций; не следует воспринимать данную схему как подробный реальный механизм осуществления терморегуляторных реакций в теле человека. Согласно этой модели температурные сигналы от различных тканей человеческого организма интегрируются на различных уровнях (включая спинной мозг и ствол мозга); при этом наиболее центром координации является гипоталамус. Вклад отдельных импульсов координируется на основе взвешенного среднего.
Регуляция температуры центральными структурами происходит путем сравнения интегрированных температурных сигналов от поверхности кожи, нервной трубки (спинного мозга и ствола мозга) и глубоких тканей с пороговыми значениями (пусковыми, или триггерными центральными температурами) для каждой отдельной терморегуляторной реакции.
Центральные контролирующие действия имеют определенную структуру: интеграция входящих сигналов осуществляется на различных уровнях нервной трубки; у млекопитающих главная роль принадлежит гипоталамусу; при этом центр вегетативной регуляции расположен в переднем гипоталамусе; а центр поведенческой регуляции – в заднем гипоталамусе. Подобная иерархическая система, по-видимому, сформировалась в процессе эволюции системы терморегуляции путем ассимиляции ранее существовавших механизмов67. Так, например, мышцы, появившиеся в процессе эволюции для поддержания позы и передвижения, были приспособлены для дрожи; аналогично, терморегуляторную вазомоцию можно рассматривать как боковую ветвь развития системы контроля гемодинамики. Вероятно, некоторые терморегуляторные реакции осуществляются, не поднимаясь выше уровня спинного мозга74. Так, у пациентов и животных с высоким пересечением спинного мозга терморегуляция становится значительно хуже; но пойкилотермными их назвать нельзя.
Угол наклона графика, отражающего взаимосвязь интенсивности ответной реакции и центральной температуры, показывает прирост терморегуляторной реакции. Максимальной интенсивностью реакции называют такую, при которой дальнейшее изменение центральной температуры уже не вызывает увеличения интенсивности реакции.. На рис.8, например, показана зависимость возникновения потоотделения от температуры, измеренной в дистальном отделе пищевода, в процессе согревания поверхности тела.
|
Рис.8. Особенности потоотделения у «среднестатистических» мужчин добровольцев демонстрируют наличие порогового значения, прироста и максимальной интенсивности во время гипертермии без использования анестезии (концентрация изофлюрана в выдыхаемом воздухе - 0%), и на фоне анестезии при концентрации изофлюрана в выдыхаемом воздухе соответственно 0,8% и 1,2%. Пороговые значения существенно увеличивались при анестезии; а прирост и уровень максимального потоотделения были практически одинаковыми во всех группах. Данные Washington et al.75; публикуется с разрешения. По вертикальной оси: потоотделение (г/м2/ч), По горизонтальной оси: пищеводная температура |
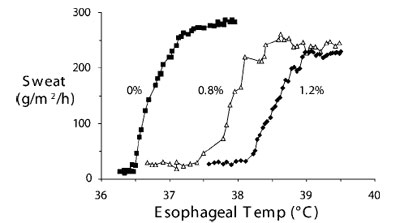
Так, пока дистальная пищеводная температура не достигнет порогового значения – 36,5°С, происходит только базовая незаметная отдача воды кожей без анестезии. Потоотделение быстро растет при увеличении центральной температуры на 0,5°С (=прирост реакции), в дальнейшем, достигнув максимума, интенсивность потоотделения остается неизменной, несмотря на дальнейшее возрастание гипертермии (=максимальная интенсивность реакции). Хотя пороговое значение для возникновения прироста меняется в зависимости от концентрации изофлюрана, скорость прироста и максимальная интенсивность остаются схожими в процессе анестезии75.
Контроль вегетативных реакций приблизительно на 80% определяется импульсами, поступающими от центральных структур76,77; и остается практически неизменным во время анестезии (рис.9). Добрая половина импульсов, определяющих поведенческие реакции, напротив, происходит с поверхности кожи78.
|
Рис.9. Индивидуальная средняя температура кожи и центральная температура при пороге вазоконстрикции (обозначено квадратами) и пороге дрожи (обозначено кружками) у 8 добровольцев. Продемонстрирована линейная зависимость между средней кожной и центральной температурами при пороге вазоконстрикции и пороге дрожи у каждого исследуемого (линии): r2=0,98±0,02 для вазоконстрикции и 0,96±0,04 для дрожи .Относительный вклад кожной и центральной температур отличался от исследуемого у исследуемому, но в среднем вклад температуры кожи составил 21±8% в вазоконстрикцию и 18±10% в дрожь. Данные Lenhardt et al.89, публикуется с разрешения. По горизонтальной оси: средняя температура кожи, по вертикальной оси: центральная температура |
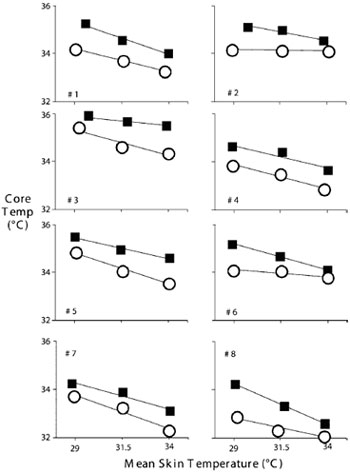
Очевидно, что человеческий организм в состоянии оценить величину собственной температуры тела с высокой точностью; тем не менее существует не конкретное значение, а некий межпороговый диапазон (набор температур), в пределах которого компенсаторные вегетатативные реакции не возникают. Диапазон температуры, которая организмом воспринимается как правильная центральная температура, определяется рядом обстоятельств (а именно, время суток, фаза менструального цикла). Нормальная центральная температура тела у людей находится в диапазоне от 36,5°С до 37,5°С; Значение центральной температуры меньше 36°С и больше 38°С, как правило, свидетельствует либо о нарушении терморегуляции, либо о чрезвычайно неблагоприятной температуре окружающей среды, при которой терморегуляционные защитные механизмы оказываются неэффективными.
Работа механизмов по поддержанию правильной схемы температуры усложняется параллельной активностью других реактивных механизмов (а именно, контроль сосудистого объема) а также факторами, зависящими от времени. В центре внимания физиологов остаются проблемы приспособления человека к выживанию к таких условиям окружающей среды, которые в норме должны запускать абсолютно противоположные механизмы компенсации. Например, тепловой удар, как правило, сопряжен с обезвоживанием в условиях очень горячей окружающей среды. Обезвоживание как таковое в принципе должно запускать механизмы, позволяющие удерживать жидкость, а гипертермия должна вызывать потоотделение. На самом деле тепловой удар обычно потому и развивается, что организм не может одновременно эффективно решить эти две задачи.
Большинство моделей терморегуляции (включая и описанную выше) недостаточно учитывают скорость изменения центральной и периферической температур. Поэтому эти модели должны применяться при динамичном развитии событий с особой осторожностью. Аналогично, существуют ситуации, когда терморегуляторные реакции определяются не только моментальными сигналами о текущей температуре, но и обусловлены предшествующими особенностями нарушений терморегуляции. Каково соотношение влияния на терморегуляторные реакции факторов, зависящих от температуры, и факторов, обусловленных временем, остается неясным.
Пороговые значения . Каким образом у человека определяются абсолютные пороговые значения температур, не вполне ясно; похоже, что при этом задействованы тормозящие постсинаптические потенциалы в гипоталамических нейронах79, работа которых регулируется адреналином, допамином, 5-гидрокситриптамином, ацетилхолином, простагландином Е и нейропептидами. Пороговые значения меняются в течение дня в диапазоне 0,5-1°С и у женщин, и у мужчин (циркадный ритм)80, и приблизительно в пределах 0,5°С у женщин в течение менструального цикла81. На величины пороговых значений также могут влиять физическая нагрузка, характер питания, наличие инфекции, гипо- и гипертиреоидизм, прием лекарственных препаратов (в том числе спирт, седативные препараты и никотин), а также холодовая и тепловая адаптация. Но влияние всех этих факторов очень мало по сравнению с глубокими нарушениями, вызываемыми общей анестезией.
Межпороговый диапазон (диапазон значений центральной температуры, при котором не запускаются регулирующие вегетативные реакции) ограничен порогом потоотделения сверху и порогом вазоконстрикции снизу. В пределах этого диапазона величина центральной температуры распознается правильно, но колебания температуры не запускают механизмы терморегуляции. С позиции целесообразности принесение в «жертву» небольшой доли терморегуляции выглядит весьма благоразумно: энергия и питательные вещества не тратятся активно для противодействия незначительным изменениям окружающей среды.
У человека внутрипороговый диапазон обычно составляет всего 0,2-0,4°С82, и это и есть диапазон нормальной температуры тела. По непонятным причинам температурный контроль в момент низшей точки циркадного ритма (примерно в 3 часа ночи) в два раза «свободнее», чем в другое время. (рис.10)80.
|
Рис.10. Межипороговый диапазон (МПД - ITR ) от потоотделения до вазоконстрикции в разные моменты дня. Результаты представлены в виде «среднее значение плюс-минус среднеквадратическое отклонение». Пороговые значения в три часа ночи значительно отличаются от результатов, полученных в любое другое время суток. Данные Tayefeh et al.80 (стр.403, рис.2) публикуется с разрешения |

У некоторых животных (верблюды, пустынные крысы) межпороговой диапазон очень широкий; колебания центральной температуры в течение дня могут достигать 10°С; это связано с наличием системы консервации энергии и питательных веществ,которая позволяет поддерживать температуру в рамках межпорогового диапазона без активного участия вегетативной нервной системы и без испарения жидкости. Но это по большому счету исключение из правил; большинство млекопитающих весьма жестко контролируют свою центральную температуру.
Оба порога – и потоотделения, и вазоконстрикции, у женщин на 0,3-0,5°С выше, чем у мужчин, даже во время фолликулярной фазы менструального цикла (т.е. в его первые 10 дней)75. Разница еще более выражена во время лютеиновой фазы83. Центральный механизм терморегуляции вполне работоспособен даже у незначительно недоношенных детей84; но, по-видимому, не полностью сформирован у глубоко недоношенных детей (весом меньше килограмма). Пороговое значение для дрожи сохраняется даже у некоторых людей в возрасте почти 90 лет, хотя это происходит и не у всех; у лиц же младше 80 лет терморегуляция практически не меняется.85
Центробежные ответные реакции . Некоторые терморегуляторные реакции весьма редко, если вообще когда-либо, активируются чем-то, кроме изменений температуры. К таким реакциям относятся потоотделение и метаболизм бурого жира. В других случаях система терморегуляции ассимилирует исполнительные механизмы, эволюционно появившиеся для других целей: это дрожь (мышечная активность для поддержания позы и направленного движения) и сосудодвигательная реакция ( контроль давления крови и осмотического статуса). Приспособление ранее созданных механизмов для нужд терморегуляции вполне согласуется с иерархической моделью терморегуляции, предложенной Satinoff67, и отчасти объясняет, почему контроль терморегуляции столь широко расходуется.
Нарушения температуры (определяемые как отклонения температуры тела от специфического порогового значения) запускают исполнительные механизмы, которые фактически содействуют соответствующему увеличению отдачи тепла в окружающую среду или увеличению метаболической продукции тепла. У каждой ответной реакции есть свой собственный порог и свой диапазон «прироста». Система контроля при этом способна к активации реакций в определенном целесообразном порядке (т.е.вазоконстрикция предшествует дрожи, более энергозатратному механизму) и в той степени, которая реально необходима для поддержания центральной температуры.
Поведенческая регуляция . Поведенческая регуляция (преднамеренное воздействие на обмен тепла с окружающей средой) является наиболее мощным исполнительным терморегуляторным механизмом. Это то новшество, которое позволяет людям существовать и в самых холодных, и в самых жарких местах на Земле. У животных поведенческий механизм регуляции температуры тоже известен. Наиболее поразительна поведенческая регуляция у рептилий и амфибий. Эти животные, которых зачастую называют «холоднокровными», на самом деле очень здорово умеют регулировать свою температуру и даже способны к поведенческой «лихорадке»86. Если их поместить в среду, диапазон температуры которой совместим с их существованием, «холоднокровные» сумеют пристроиться так, чтобы поддерживать центральную температуру в пределах нескольких градусов от «нормальной». Любопытно, что оптимальной для большинства рептилий является температура приблизительно такая же, как и у млекопитающих, около 37°С. Аналогичным образом поступают рыбы87. Один исследователь даже смог «надрессировать» свою золотую рыбку поддерживать нужную температуру воды (а, следовательно, и своего тела) путем нажатия кнопки88. Даже бактерии при предоставлении такой возможности будут стремиться расположиться там, где есть возможность поддерживать оптимальную температуру.
Активное поведенческое изменение отдачи тепла в окружающую среду как правило не требуется, если температура окружающей среды приемлема. В этом заключается эволюционное достижение - поддержания практически постоянной центральной температуры (вероятно необходимой для оптимального функционирования ферментов) без использования поведенческих реакций, которые могут угрожать выживанию. Тем не менее, если вегетативных реакций терморегуляции оказывается недостаточно для поддержания центральной температуры, принципиальными для выживания становятся поведенческие механизмы. Поведенческое приспособление очень разнообразно; из наиболее распространенных вариантов можно отметить столь простые действия как уход с прямого солнца в тень, выбор одежды по погоде, использование кондиционеров и отопления. Поведенческие реакции требуют сознательного ощущения температуры тела. Интересно, что человек плохо ощущает изменения центральной температуры; и напротив, очень чутко чувствует даже минутные колебания температуры поверхности кожи. Таким образом, поведенческая терморегуляция приблизительно наполовину обусловлена температурой кожи78, тогда как в работе вегетативных терморегуляторных механизмов доля информации, поставляемая кожей, составляет всего лишь 10-20%76,89.
Сосудо-двигательные реакции . Отдача метаболического тепла преимущественно производится через поверхность кожи, и сужение сосудов кожи, наиболее часто используемый исполнительный механизм вегетативной терморегуляции, уменьшает потерю тепла. В суммарном кожном токе крови можно выделить питающую (в основном капилляры) и терморегуляторную (в основном артериовенозные шунты) составляющие90. Шунты обычно имеют диаметр 100 μm; это означает, что один шунт может передать в 10000 раз больше крови, чем соответствующей длины капилляр 10 μm в диаметре. Артериовенозные шунты могут быть открытыми и закрытыми, что собственно означает большой возможный диапазон «прироста» при использовании данного механизма терморегуляции. Порядка 10% сердечного выброса проходит через артериовенозные шунты; соответственно, «закрытие» шунтов увеличивает артериальное давление приблизительно на 15 мм рт ст91.
Артериовенозные шунты располагаются только в дистальных отделах (пальцы рук, пальца ног, нос и т.д.) Эти специализированные терморегуляторные сосуды являются α-адренергическими; их сужение происходит под действием норадреналина, выделяющегося симпатическими нервными окончаниями. Похоже, что циркулирующие в крови вещества не оказывают прямого влияния на артериовенозные шунты, хотя гормоны (такие как ангиотензин) могут способствовать ответу на полученный симпатический стимул. Большинство сосудов при локальной гипотермии сужаются, но артериоввенозные шунты относительно устойчивы к местным температурным изменениям и, скорей всего, контролируются исключительно центральным терморегуляторным статусом. При термонейтральной окружающей среде (т.е.температура воздуха соответствует температуре тела в пределах межпорогового диапазона) и в денервированной конечности артериовенозные шунты полностью открыты. Впрочем, при обычной температуре окружающей среды тонизирующая симпатическая симуляция поддерживает кровоток в шунтах на минимальном уровне.
« Несократительный» термогенез . Под несократительным термогенезом понимают увеличение метаболической продукции тепла, несвязанное с работой мышц. Такое увеличение преимущественно происходит в специализированной жировой ткани - буром жире, располагающемся в основном между лопатками и около почек. Бурый жир имеет коричневый оттенок за счет чрезвычайно высокого содержания митохондрий. При стимуляции бурый жир обладаем самой высокой метаболической активностью в человеческом организме (до 0,5 Вт/г). Обычно, митохондриальный метаболизм приводит к образованию протона, который секретируется за пределы саркоплазматической сети. Протонный трансмембранный градиент в свою очередь активирует калий-натриевую АТФ-азу, приводя к образованию АТФ из АДФ. При высвобождении норадреналина из симпатических нервных окончаний митохондриальное дыхание в бурой аденозин- трифосфатазной ткани происходит обычным образом. При этом продукции АТФ препятствует специальный «расщепляющий белок», который позволяет протонам вернуться в саркоплазматическую сеть без использования калий-натриевой АТФазы92.
Несократительный термогенез является основным механизмом защиты от холода у мелких млекопитающих (мыши, крысы); он может с легкостью приводить к удвоению, а то и утроению метаболической продукции тепла (оцениваемой по потреблению организмом кислорода) без выполнения какой-либо механической работы. Недрожательный термогенез может удваивать продукцию тепла и у младенцев93. Интенсивность недрожательного термогенеза является линейной функцией от разницы между величиной средней температурой тела (СТТ) и ее пороговым значением.
Несократительный термогенез имеет важное значение только у маленьких животных и у младенцев; при относительно крупных размерах биологического организма (>50кг) этот механизм или не имеет реального практического значения, или не работает вообще. У взрослых людей такой термогенез слабо развит и мало участвует в поддержании температуры тела
Дрожь . Длительная дрожь увеличивает метаболическую продукцию тепла у взрослых на 50-100%. Это увеличение очень мало в сравнении с увеличением, происходящим вследствие физической нагрузки (при физической нагрузке увеличение продукции тепла может быть пятикратным, по крайней мере, кратковременно), и поэтому является на удивление малоэффективным. Дрожь это беспорядочное сотрясение, которое выглядит на электромиограмме как случайные накладывающиеся зубцы деполяризации миофибрилл. На эти быстрые и явно хаотические всплески местной активности накладываются более структурированные волнообразные циклы изменения активности (4-10 колебаний в минуту). Причем, эти медленные волновые изменения активности происходят синхронно во всех мышцах тела95. Дрожание не встречается у новорожденных младенцев и скорей всего недостаточно эффективно у детей первых лет жизни. Так как температурный порог дрожи на целый градус меньше порога вазоконстрикции82, похоже, что это «последняя надежда» среди способов борьбы с переохлаждением.
Потоотделение . Потоотделение контролируется постганглионарными холинергическими нервными волокнами. Это активный процесс, который может быть выключен посредством нервной блокады или введением атропина97. Даже у нетренированных людей потоотделение может достигать до 1 л/ч, а спортсмены могут «потеть» в два раза интенсивнее. Потоотделение является единственным механизмом отдачи тепла, работающим при температуре окружающей среды большей, чем центральная температура тела. К счастью, этот механизм работает очень эффективно: Вместе с одним граммом пота «уходит» 0,58 ккал. Если окружающий человека воздух сухой и есть конвекция, величина теплоотдачи путем потоотделения может во много раз превышать значение базального метаболизма, который в грубом приближении составляет ккал/кгхчас. Конечно, пот, который образуется скапливается на коже в виде капелек и не испаряется, никак не способствует тепловому балансу и лишь содействует обезвоживанию.
При физической нагрузке мышечный кровоток резко увеличивается, и давление крови можно поддержать только за счет вазоконстрикции в других сосудистых сетях. Более того, при физической нагрузке образуется столько тепла, что для его эффективной отдачи практически при любой погоде требуется усиление капиллярного кровотока и потоотделения (1 л/ч или даже больше). При этом механизмы терморегуляции вступают в «борьбу» с потребностью мышц в увеличении кровотока. Так что неудивительно, что максимальный капиллярный кровоток и интенсивность потоотделения могут повреждаться при снижении ОЦК и сердечно-сосудистой недостаточности. Учитывая серьезное стрессовое действие физической нагрузки на сердечно-сосудистую систему и терморегуляторную компенсацию возникающего при этом увеличения метаболической продукции тепла, поразительно, как вообще удается человеку вести себя энергично при высокой температуре окружающей среды и сохранять приемлемый уровень артериального давления.
В противоположность кровотоку в шунтах, капиллярный кровоток минимален и при обычных температуре окружающей среды, и при термонейтральном окружении. При тепловом стрессе активная дилятация прекапиллярных артериол приводит к резкому возрастанию капиллярного кровотока. Эта дилятация безусловно связана с прекращением симпатической стимуляции, но, похоже, что в возникновении дилятации какую-то роль играет и какое-то химическое вещество, продуцируемое потовыми железами; что это за вещество, еще предстоит определиться, возможно, это окись азота или нейропептид Y98. Так как активная вазодилатация требует сохранной функции потовых желез, она также значительно угнетается при нарушении нервной передачи. При серьезном тепловом стрессе кровоток в поверхностном слое кожи толщиной 1 мм может достигать 7,5 л/мин – то есть столько же, сколько составляет весь сердечный выброс в покое99. Пороговое значение для активной вазодилатации аналогично пороговому значению потоотделения, но обычно максимальная кожная вазодилатация запаздывает и наступает после достижения максимальной активности потоотделения.
Стратегия активации ответных реакций . В идеальном варианте все потенциальные терморегуляторные реакции доступны и используются в определенном порядке в зависимости от порогового значения и диапазона ответных приростов. Однако при определенных обстоятельствах один или несколько ответных механизмов могут оказаться недейственными. Например, социальные обычаи могут ограничивать свободу произвольного передвижения или возможность искать более теплое или прохладное место. Наличие в организме миорелаксанта может препятствовать дрожи, а вазодилататора – ваконстрикции. В таких ситуациях механизмы, оставшиеся рабочими, пытаются справиться с ситуацией своими силами и компенсировать недостающие механизмы в пределах своих возможностей. Как правило, при этом центральную температуру удается удержать, но сокращаются границы возможности приспособления к состоянию окружающей среды.
Фармакогенетические аспекты клинической анестезиологии А. В. Бурлев, Е. М. Шифман ФГУ Научный центр акушерства гинекологии и перинатологии им. акад. В. И. Кулакова Минздравсоцразвития РФ, Москва |
Настоящий обзор литературы с современных позиций раскрывает вопросы фармакогенетики и фармакогеномики, номенклатуры полиморфизмов, основные клинически значимые полиморфизмы для наиболее распространенных в клинической анестезиологии классов препаратов. Проанализированы 53 источника литературы, а также база данных полиморфизмов Национального центра биотехнологической информации (США). Сегодня выбор препарата не предусматривает межличностную генетическую вариабельность. Вполне вероятно, что определенные препараты обладают потенциалом для развития серьезных побочных эффектов или являются неэффективными у определенного подкласса популяции вследствие неблагоприятного соотношения эффективность/токсичность. С прогрессом технологии, каждое генетическое подмножество популяции вероятно, будет пролечено специфически полученным препаратом для обеспечения искомого эффекта. Возможно еще рано ожидать непосредственных результатов фармакогенетики для повседневной практики анестезиолога, вероятно, что прогресс в области геномики даст лучшее понимание SNPs и гаплотипов, которые изменяют реакции на препарат. Эти открытия в будущем позволят "нацеливание"анестезиологических препаратов и персонализацию стратегий обезболивания, направленных на улучшение периоперационных исходов, а также на лечение острой и хронической боли, в соответствии с генетическим профилем наших пациентов.
Ключевые слова: фармакогенетики, фармакогеномика, однонуклеотидный полиморфизм, анестезиология
Введение. Фармакогенетика внедрилась в сферу анестезиологии в середине 50-х годов XX века с появлением первых наблюдений, свидетельствующих, что генетические факторы человека могут изменять реакцию на различные фармакологические препараты. Первые статьи в этой области сообщали о случаях пролонгированной послеоперационной мышечной релаксации вследствие вариации бутирилхолинэстеразного пути метаболизма сукцинилхолина [1]. Понятие фармакогенетика и фармакогеномика используют как синонимы, но концепцию фармакогеномики можно представить как более "масштабную", включающую анализ множества генетических процессов и путей одновременно, а не друг за другом, а также исследования для определения унаследованной реакции на препарат и геномные тесты. Около 90% генетических вариаций возможно в результате появления так называемых однонуклеотидных полиморфизмов. Если несколько полиморфизмов происходят в одном и том же гене, то результирующий генотип называется "гаплотип" [2].
Однонуклеотидный полиморфизм (англ. Single nucleotide polymorphism — SNP) — отличия последовательности ДНК размером в один нуклеотид (А, Т, G или С) в геноме (или в другой сравниваемой последовательности) представителей одного вида или между гомологичными участками гомологичных хромосом индивида. Частота встречаемости составляет 1 SNP на 100—300 базовых пар или "букв" генома человека. В настоящее время подробный учет и картирование осуществляется Национальным центром биотехнологической информации (США) и на момент написания обзора последнее обновление базы данных полиморфизмов датировалось 1 апреля 2010 г. (версия db-SNP 131) [3].
Учитывая, что только от 3 до 5% ДНК-последовательностей индивида кодируют продукцию белка, большинство полиморфизмов находятся вне кодируемой области. Однако полиморфизмы, находящиеся внутри кодируемой области генов, способны в значительной степени влиять на укладку протеина в спиральную структуру, смену полярности, приводить к ошибкам фосфорилирования и как следствие изменять биологические функции белка [4].