

Введение
В прошлом, методы для оценки эффектов анестезии на активность головного мозга были основаны только на измерении поведенческих исходов или на электрофизиологических данных, таких как запись электроэнцефалограмм ( EEG ) или вызванных потенциалов [1]. В прошлой декаде прогресс в нейро-научных методов расширил этот предел методов, включив позитронную эмиссионную томографию ( PET ) и функциональное изображение магнитного резонанса ( fMRI ) [2]. Эти методы определяют место гемодинамических и метаболических изменений, вызванных анестезия-индуцированными изменениями активности головного мозга, в то время как электрофизиологические методы отражают электрическую активность коры с высокой временной, но низкой пространственной разрешающей способностью.
В клиническом образце, текущие разработки нейромониторинга (например, биспектрального индекса ( BIS )) позволяют дать общую оценку нейронной активности во время операции [3]. Эти методы, однако, не прогнозируют движения или гемодинамические реакции на стимуляцию, но они не могут прогнозировать точно, когда отдельные пациенты будут восстанавливать сознание [4]. Отсюда, информированность (т.е. эксплицитная память) [5] и формирование имплицитной памяти наблюдается [6] даже при предположительно адекватной анестезии, руководствуясь нейромониторингом. Таким образом, важные аспекты церебральной функциональности во время анестезии остаются неизвестными [7]. С помощью дополнительных возможностей электрофизиологических методов и методов изображения мозга, этот пробел отчасти закрыт в современные годы при исследовании эффектов препаратов на нейронные связи, вовлекаемые в формирование внимания, процесс слуховой обработки данных, памяти и сознания. Более того, детальные исследования с четким контролем гипнотического состояния, адресованные бессознательному формированию памяти, уже выполнены. Таким образом, этот обзор суммирует современные данные, полученные различными методами исследования, и представляет систему взглядов, которая интегрирует текущую информацию.
Активность головного мозга во время анестезии, измеренная с помощью функционального нейроизображения
Функциональное нейроизображение - это в основном современные методы изображения головного мозга, такие как PET и fMRI . Оба метода успешно применяются в фармакологических исследованиях и при исследовании действия анестетиков в последнее десятилетие.
Изменения церебрального кровотока , церебрального метаболизма и оксигенации крови уровень-зависимым контрастом
Исследования PET и fMRI показывают изменения активности мозга вследствие действия анестетиков, при условии, что сохраняются нейроваскулярные связи при анестезии [8]. С помощью этих методов, доза-зависимое снижение церебрального кровотока ( CBF ), церебрального метаболизма глюкозы и оксигенации крови доза-зависимым ( BOLD ) сигналом показано примерно для всех анестетиков [9-14,15,16], за исключением кетамина [17,18] и закиси азота [19]. Эти действия указывают на общее снижение нейронной активности при углублении анестезии. Эффективность при снижении активности головного мозга предположительно одинакова для внутривенных и ингаляционных анестетиков, что оценивались относительно схожими снижениями церебрального метаболизма на сопоставимых уровнях анестезии севофлюраном и пропофолом [19]. Потеря сознания происходит при церебральном метаболизме глюкозы примерно 60-65% [20]. За исключением общего снижения нейронной активности, некоторые области головного мозга, расположенные в пределах ассоциативных полей коры головного мозга (например, теменная и лобная ассоциативная кора), таламусе и в среднем мозге, показывают заметно более выраженное снижение церебрального кровотока, церебрального метаболизма или сигнала BOLD при сравнении с общим снижением [9-14, 15, 16, 19, 21-24] (смотри таблицу 1). Эти действия показывают специфические эффекты анестетиков для создания бессознательного состояния, амнезии и нарушения внимания: существуют убедительные доказательства, что таламус играет ключевую роль при анестезия-индуцированном бессознательном состоянии [25, 26], так как относительное метаболическое снижение регионального метаболизма глюкозы [12, 25] и CBF [27] наблюдалось в этой области при адекватной анестезии. И наоборот, низкие концентрации анестетиков, как оказалось, не оказывают или оказывают только небольшой эффект на таламус [22] и области, вовлеченные в первичную обработку информации (рис. 1 и 2).
|
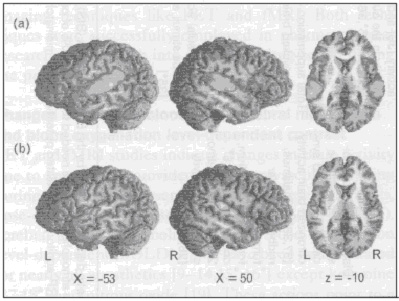
Рис 1. Активность мозга во время обработки лингвистической информации (бодрствование против седации). Функциональная активность головного мозга, вызванная слуховой обработкой данных, полученной во время бодрствования ( a ) и во время глубокой седации (т.е., когда индивидуумы отвечают на задачи языкового процесса) ( b ). Во время бодрствования, слуховая обработка данных вызывает активность мозга в височно-лобной связи. Глубокая седация (пропофол эффективная местная концентрация в пределах между 1,51 и 3,35 мкг/мл) ограничивает измеренную активность мозга в областях в и вокруг первичной слуховой коры в обоих полушариях. При эффективных местных концентрациях пропофола, превышающих 3,35 мкг/мл, не выявлена функциональная активация мозга, связанная со слуховой стимуляцией. Изменено с разрешения Heinke et al . [15]. |
|
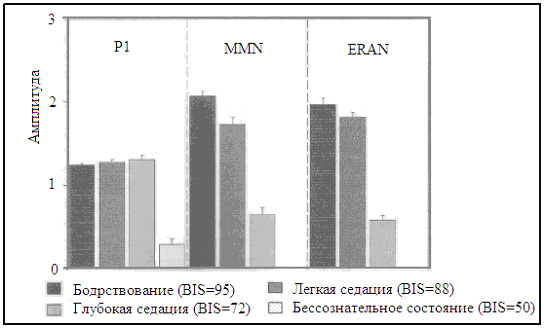
Рис 2. Эффект повышенной седации пропофолом на слуховые потенциалы. Отражает первичный процесс обработки слуховой информации ( P 1), слуховую сенсорную память (MMN) и музыкальный синтаксический процесс обработки данных (ERAN). Амплитуда (средняя величина ± среднеквадратическая ошибка) P1, MMN и ERAN во время различных уровней седации, урегулированная с целе-контролируемой инфузией пропофола: бодрствование (концентрация пропофола 0,0 мкг/мл, средний биспектральный индекс (BIS = 95), легкая седация (концентрация пропофола 0,5 мкг/мл, средний BIS = 89), глубокая седация (концентрация пропофола 1,5 мкг/мл, средний BIS = 72) и бессознательное состояние (концентрация пропофола 2,5-3,0 мкг/мл, средний BIS = 50). Амплитуда P 1 не изменялась при седации и заметно снижалась, но определялась, при бессознательном состоянии. И наоборот, амплитуда MMN и ERAN постепенно снижалась при повышении седации и исчезала при бессознательном состоянии. Это показывает характерные эффекты седации пропофолом на слуховую сенсорную память и музыкальный синтаксический процесс обработки данных при сравнении с первичным процессом звукового входа информации. Изменено с разрешения Heinke et al. [36] (смотри ссылки для получения дальнейших деталей). |
Низкие дозы препаратов, однако, как предполагается, вызывают амнезию либо посредством относительно неспецифической депрессии нейронной активности преимущественно в коре [28] или специфическими успокоительными эффектами на гипокамп [29], островковую долю большого мозга [22, 30], мозжечковую миндалину [31] и предлобную кору [30]. Эти действия анестетиков, как оказалось, поражают память путем нарушения кодирования новой информации в память (неспецифический эффект препарата вследствие седации) и путем нарушенного сохранения кодированного материла в долговременную память. Последний эффект, как, оказалось, является независимым от седации и вероятно отражает специфическое амнестическое действие анестетиков [32].
Хотя эти данные отчасти объясняют бессознательное состояние и амнезию, они не обеспечивают никакой информацией о нейронной активности, связанной с сенсорной стимуляцией во время анестезии. Исследования на животных предполагают, что спровоцированная активность коры может быть выявлена даже во время глубокой анестезии [33]. У человека, нейронная активность уже отмечалась с помощью функционального нейроизображения, вызванного другими стимуляторами [15, 30, 34], зрительным [13] или вредными стимулами [35] при поверхностной и адекватной стадиях анестезии (таблица 1). Например, мы наблюдали функциональную активность в обеих височных долях вызванную слуховым процессом обработки данных при эффективной местной концентрации пропофола выше 1,51 мкг/мл (рис. 1). Эта активация исчезала при концентрации, превышающей 3,35 мкг/мл. Подобным образом, севофлюран снижал слуховую спровоцированную функциональную активацию по доза-зависимому манеру. При сравнении с бодрствованием, остаточная активация была отмечена с двух сторон в верхней височной извилине, таламусе, стриатуме и левой лобной коре при 1% севофлюране, в то время как эта активность полностью подавлялась при 2% севофлюране [34]. В настоящее время, показано, что седативные дозы пропофода и тиопентала не изменяют повышения CBF , вызванного слуховыми стимулами, несмотря на 15% снижение общего CBF (gCBF) [41]. Даже при невосприимчивости (средний BIS = 66), реакция регионального CBF ( rCBF ) в левой височной доле была отмечена (хотя явно сниженная). В целом, эти данные предполагают, что слуховая стимуляция все еще вызывает активность коры, хотя и сниженную, во время поверхностных стадий анестезии.
Суммарно: анестезирующее действие на активность мозга в покое, спровоцированная активность мозга и когнитивные функции в зависимости от глубины анестезии. Глубина анестезии показана по BIS , эффекту местной концентрации пропофола и минимальной альвеолярной концентрации ( MAC - это минимальная альвеолярная концентрация ингаляционных анестетиков при давлении в одну атмосферу, необходимая для предотвращения движения в ответ на хирургический разрез кожи у 50% пациентов). gCBF – общий церебральный кровоток; gCMR glu – общая церебральная скорость метаболизма глюкозы; aERP – слуховой спровоцированный потенциал; MMN - неподходящая отрицательность; ERAN – ранняя правая передняя отрицательность .
Подобно слуховым стимулам, соматосенсорные стимулы могут также вызывать активность коры во время невосприимчивости, но это вероятно зависит от использованных стимулов. Вибротактильные стимулы не активируют кору и вызывают только активность таламуса при концентрации пропофола 1,5 мкг/мл [39]. И наоборот, вредные (жар) стимулы повышают CBF в соматосенсорной и средне-островковой коре даже при концентрации пропофола 3,5 мкг/мл (т.е., во время бессознательного состояния) [35]. Интересно, что передняя поясная кора – область, включающаяся в болевое восприятие [42], активируется жарой при более низких концентрациях пропофола. Таким образом, островковая доля большого мозга может быть активирована даже во время бессознательного состояния (хотя не вызывает болевого восприятия, а это скорее автономные реакции), в то время как другие структуры, отвечающие за болевое восприятие, оказались подавленными при концентрациях пропофола, вызывающих бессознательное состояние [35].
Эффекты кетамина на CBF [17] и церебральный метаболизм глюкозы [17, 18, 43] при активности мозга в покое ясно характеризуется для других анестетиках. Субанестезирующие дозы кетамина вызывают общее повышение rCBF , наиболее глубокие изменения в структурах головного мозга, связанных с процессом обработки боли [17]. Под вредной стимуляцией, однако, субанестезирующие дозы кетамина снижают болевые и стимулы, вызывающие активность мозга, с наибольшим снижением, наблюдаемым в таламусе и островковой доле большого мозга [44]. Помимо этих эффектов обезболивания, кетамин, как оказалось, в низких дозах влияет на активность головного мозга, связанную с «рабочей» памятью, в областях мозга, включающих лобную и теменную области коры и скорлупу [45], а также на активность, связанную с кодированием и восстановлением случайной информации в лобной и в гиппокампальной областях [46].