

ekzamen_po_mikre_udacha
.pdfБольшинство прокариот имеет ригидную клеточную стенку, под которой расположена ЦПМ. Состав и строение клеточной стенки — важный систематический признак, по которому все прокариоты подразделяются на следующие группы:
Гр+ бактерии |
Бактерии |
Грбактерии |
|
не |
|
|
имеющие |
|
|
клеточной |
|
|
стенки |
|
Клеточная стенка содержит до 40 слоев пептидогликана. Молекулы тейхоевых кислот ковалентно связаны с пептидогликаном. Липотейхоевые кислоты – тейхоевые кислоты, содержащие липидные «хвостики», которые закреплены в гидрофобной области ЦПМ. Клеточные стенки могут иметь белковые слои на поверхности. Белковые структуры могут быть расположены островками, или быть тесно упакованными и образовывать S-слой.
Грамотрицательные бактерии обладают более сложной клеточной оболочкой, имеющей внешнюю мембрану, в которой внутренний слой состоит из фосфолипидов, а внешний
— из липополисахаридов. Между цитоплазматической и внешней мембранами возникает уникальное образование, называемое периплазматическим пространством (периплазмой). Муреиновый мешок таких бактерий встроен в периплазму и состоит всего лишь из одного слоя (толщина ~3 нм). ВМ и муреиновый слой соединены липопротеином. ВМ содержит белки-порины, формирующие поры.
Бактерии обычно имеют ригидную клеточную стенку, основным опорным элементом которой является муреин — поперечносшитый биополимер, гетерополисахарид, формирующий замкнутый мешок, полностью покрывающий клетку снаружи. Муреиновый мешок не только стабилизирует клеточную мембрану, но и сохраняет неизменной форму клетки.
 ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
располагается под клеточной стенкой (между ними - периплазматическое пространство). По строению является сложным липидобелковым комплексом, таким же, как у клеток эукариот (универсальная мембрана).
мембранные липиды образуют бислой, где неполярные области их молекул обращены друг к другу в центральной части мембраны, а их полярные группы смотрят наружу
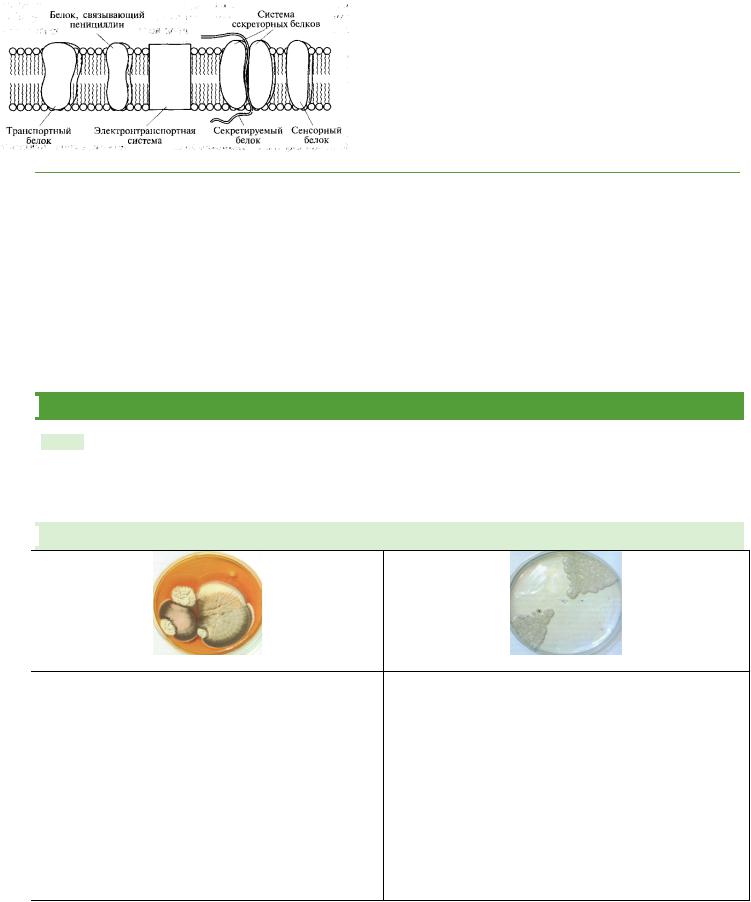
Мембранные белки пронизывают бислой мембраны и могут диффундировать в жидком матриксе, образуя иногда большие сложные комплексы.
 ФУНКЦИИ ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ:
ФУНКЦИИ ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ:
1.ЦПМ представляет собой осмотический барьер клетки
2.Осуществляет активный перенос веществ против градиента концентрации
3.Энергетическая функция
4.Биосинтетическая функция: на мембране осуществляется синтез веществ клеточной стенки
5.Секреторная функция: проведение и выделение веществ
6.Компартментация биохимических процессов
7.Интеграция биохимических процессов
8.Возбудимый акцептор и проводник сигналов
9.Участие в делении бактериальной клетки
 10. МОРФОЛОГИЯ ГРИБОВ.
10. МОРФОЛОГИЯ ГРИБОВ.
ГРИБЫ – эукариоты. Они имеют хорошо оформленное ядро, митохондрии и вакуоли. Грибная клетка содержит одно или несколько ядер. Клеточная стенка грибов состоит на 80-90% из полисахаридов (45% приходится на хитин) и 10-20% приходится на белки и липиды. Соотношение химических ингредиентов клеточной стенки различных грибов определяет их вирулентность. Строение клеточной стенки определяет выбор антибиотических средств.
 РАЗЛИЧАЮТ 2 ОСНОВНЫХ ТИПА ГРИБОВ:
РАЗЛИЧАЮТ 2 ОСНОВНЫХ ТИПА ГРИБОВ:
Колонии гифальных (плесневых) грибов
образуют ветвящиеся тонкие нити (гифы), сплетающиеся в
грибницу, или мицелий (плесень). Гифы, врастающие в питательный субстрат, называются вегетативными гифами (отвечают за питание гриба), а растущие над поверхностью субстрата — воздушными или репродуктивными гифами (отвечают за бесполое размножение).
Гифы низших грибов не имеют перегородок. Они представлены многоядерными клетками и называются ценоцитными.
Гифы высших грибов разделены перегородками, или септами с отверстиями.
Колонии дрожжевых грибов в основном, имеют вид отдельных овальных клеток
(одноклеточные грибы). По типу полового размножения они распределены среди высших грибов — аскомицет и базидиомицет.
При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту.
Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток — «сарделек».
Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом — почкованием или делением.

 11. МОРФОЛОГИЯ ПРОСТЕЙШИХ.
11. МОРФОЛОГИЯ ПРОСТЕЙШИХ.
ПРОСТЕЙШИЕ — эукариотические одноклеточные микроорганизмы, составляющие подцарство Protozoa царства животных (Animalia). Простейшие включают 7 типов, из которых четыре типа (Sarcomastigophora, Apicomplexa, Ciliophora, Microspora) имеют представителей, вызывающих заболевания у человека. Размеры простейших колеблются в среднем от 5 до 30 мкм.
•Снаружи окружены мембраной (пелликулой) — аналогом цитоплазматической мембраны клеток животных. Некоторые простейшие имеют опорные фибриллы.
•Цитоплазма и ядро соответствуют по строению эукариотическим клеткам: цитоплазма состоит из эндоплазматического ретикулума, митохондрий, лизосом, многочисленных рибосом и др.; ядро имеет ядрышко и ядерную оболочку.
•Передвигаются простейшие посредством жгутиков, ресничек и путем образования псевдоподий.
могут питаться в результате фагоцитоза или образования особых структур. Многие простейшие при неблагоприятных условиях образуют цисты — покоящиеся стадии, устойчивые к изменению температуры, влажности и др.
Простейшие окрашиваются по Романовскому—Гимзе (ядро — красного, цитоплазма — синего цвета).
 12. ОСОБЕННОСТИ МОРФОЛОГИИ ВИРУСОВ.
12. ОСОБЕННОСТИ МОРФОЛОГИИ ВИРУСОВ.
ВИРУСЫ - это мельчайшие микроорганизмы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК).
•отличаются разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки и затем происходит их сборка в вирусные частицы.
•Вирусы размножаются в цитоплазме или ядре клетки.
Сформированная вирусная частица называется ВИРИОНОМ.
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
ФОРМА ВИРИОНОВ МОЖЕТ БЫТЬ РАЗЛИЧНОЙ:
•палочковидной (вирус табачной мозаики),
•пулевидной (вирус бешенства),
•сферической (вирусы полиомиелита, ВИЧ),
•в виде сперматозоида (многие бактериофаги).
РАЗМЕРЫ:
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования.
•Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм),
•наиболее крупным – натуральной оспы (около 350 нм).
ВИРУСЫ ИМЕЮТ УНИКАЛЬНЫЙ ГЕНОМ
•ДНК-СОДЕРЖАЩИЕ или
•РНКСОДЕРЖАЩИЕ ВИРУСЫ
Они обычно гаплоидны, т.е. имеют один набор генов.
Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными.
 СРЕДИ РНК-СОДЕРЖАЩИХ ВИРУСОВ РАЗЛИЧАЮТ ВИРУСЫ:
СРЕДИ РНК-СОДЕРЖАЩИХ ВИРУСОВ РАЗЛИЧАЮТ ВИРУСЫ:

-С ПОЛОЖИТЕЛЬНЫМ (ПЛЮСНИТЬ РНК) ГЕНОМОМ. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (ИРНК).
-РНК-СОДЕРЖАЩИЕ ВИРУСЫ С ОТРИЦАТЕЛЬНЫМ (МИНУС-НИТЬ РНК) ГЕНОМОМ.
Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Геном вирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (вирусы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
РАЗЛИЧАЮТ:
- ПРОСТО УСТРОЕННЫЕ (например, вирус полиомиелита)
У просто устроенных вирусов нуклеиновая кислота связана с белковой оболочкой, капсидом (от лат. сарsа футляр). Капсид состоит из повторяющихся морфологических субъединиц и взаимодействуя друг с другом, образуют нуклеокапсид.
- СЛОЖНО УСТРОЕННЫЕ (например, вирусы гриппа, кори) вирусы.
У сложно устроенных вирусов капсид окружен дополнительной липопротеидной оболочкой структур клетки-хозяина), имеющей «шипы».
 ТИП СИММЕТРИИ КАПСИДА:
ТИП СИММЕТРИИ КАПСИДА:
Для вирионов характерен:
- СПИРАЛЬНЫЙ - обусловлен винтообразной структурой нуклеокапсида
- КУБИЧЕСКИЙ - образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту.
- СЛОЖНЫЙ
Капсид и суперкапсид защищают вирионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуногенные свойства вирионов. Внутренние структуры вирусов называются сердцевиной
 13. СТРУКТУРА И ХИМИЧЕСКИЙ СОСТАВ БАКТЕРИОФАГОВ.
13. СТРУКТУРА И ХИМИЧЕСКИЙ СОСТАВ БАКТЕРИОФАГОВ.
БАКТЕРИОФАГИ – вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродукцироваться в них и вызывать их растворение (лизис).
Фаги различаются по форме, структурной организации, типу нуклеиновой кислоты и характеру взаимодействия с микробной клетокой
СТРУКТУРА
Под микроскопом имеют форму головастика или сперматозоида, некоторые – кубическую и нитевидную формы. Размеры фагов колеблются от 20 до 800 нм у нитевидных фагов.
КРУПНЫЕ ФАГИ, ИМЕЮЩИЕ ФОРМУ СПЕРМАТОЗОИДА:
•состоят из вытянутой икосаэдрической головки размеров 65-100 нм и хвостового отростка длинной более 100 нм.
•Внутри хвостового отростка имеется полый цилиндрический стержень, сообщающийся отверстием с головкой, снаружи – чехол, способный к сокращению наподобие мышцы.
• Хвостовой отросток заканчивается шестиугольной базальной пластинкой с короткими шипами, от которых отходят нитевидные структуры – фибриллы.
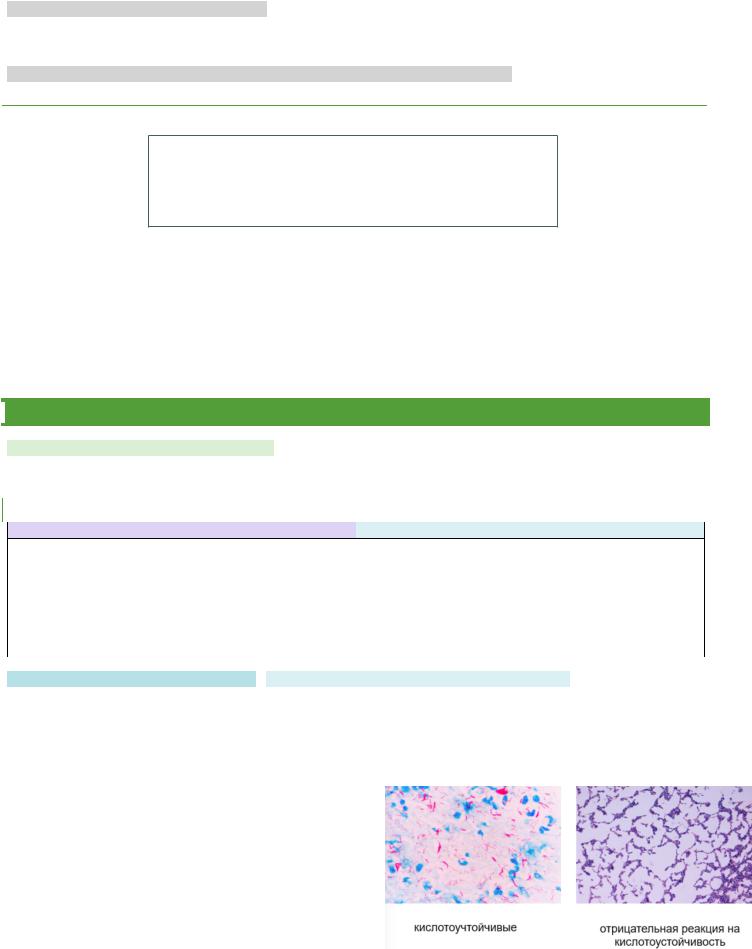
ФАГИ, ИМЕЮЩИЕ ДЛИННЫЙ ОТРОСТОК:
•Чехол не способен сокращаться
ФАГИ С КОРОТКИМИ ОТОСТКАМИ, АНАЛОГАМИ ОТРОСТКОВ, БЕЗ ОТРОСТКОВ.
 ХИМИЧЕСКИЙ СОСТАВ
ХИМИЧЕСКИЙ СОСТАВ
Состоят из 2-ух основных химических компонентов:
1)нуклеиновой кислоты (ДНК и РНК)
2)белка
*У фагов, имеющие форму сперматозоида, двунитчатая ДНК плотно упакована в виде спирали внутри головки
•белки входят в состав оболочки (капсида), окружающей нуклеиновую кислоту, и во все структурные элементы хвостового отростка.
•структурные белки фага различаются по составу полипептидов и представлены в виде множества идентичных субъединиц, уложенных по спиральному или кубическому типу симметрии.
•Кроме структурных белков обнаружены внутренние (геномные) белки, связанные с нуклеиновой кислотой, и белки-ферменты (лизоцим, АТФ-фаза), участвующие во взаимодействии фага с клеткой.
 14. ТИНКТОРИАЛЬНЫЕ СВОЙСТВА БАКТЕРИЙ. МЕТОДЫ ОКРАСКИ.
14. ТИНКТОРИАЛЬНЫЕ СВОЙСТВА БАКТЕРИЙ. МЕТОДЫ ОКРАСКИ.
ТИНКТОРИАЛЬНЫЕ СВОЙСТВА БАКТЕРИЙ – способность бактерий и их отдельных компонентов воспринимать красители
МЕТОДЫ ОКРАСКИ
ПРОСТЫЕ МЕТОДЫ |
|
СЛОЖНЫЕ МЕТОДЫ |
Один краситель (Пример: фуксин, метиленовый синий) |
На мазок последовательно наносят красители (несколько |
|
|
красителей) |
|
|
• Окраска по методу Циля-Нильсона |
|
|
• |
По Бурри-Гинса |
|
• |
По Нейссера |
|
• |
По Ожешки |
|
• |
По Граму |
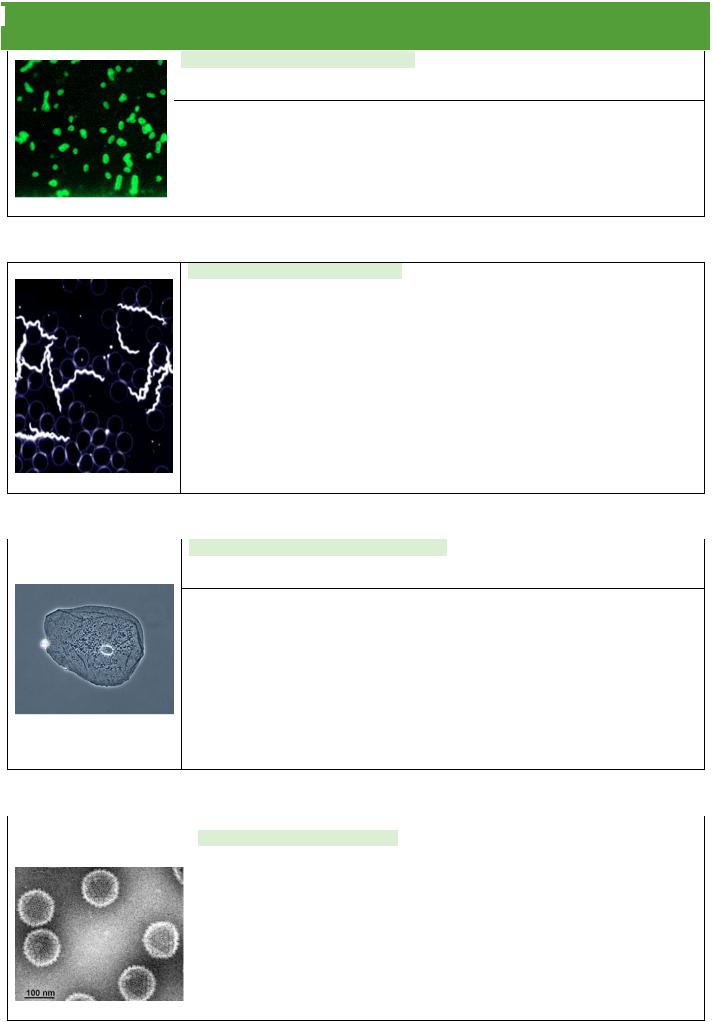
ОКРАСКА ПО МЕТОДУ ЦИЛЯ-НИЛЬСОНА– для выявления кислоустойчивости микроорганизмов.
При правильном окрашивании кислотоустойчивые бактерии приобретают рубиново-красный цвет, а все остальные синий.
•На фиксированный мазок кладут фильтровальную бумагу пропитанную раствором карболового фуксина Циля и подогревают над пламенем горелки троекратно до появления паров, держа высоко над пламенем.
•Препарату остужают, снимают фильтровальную бумагу и промывают водой.
•Затем препарат обесцвечивают 5% раствором серной кислоты, погружая предметное стекло 2-3 раза в стакан
скислотой.
•Препарат промывают водой.
•Докрашивают метиленовым синим в течение 3-5 минут.
•Краску сливают и промывают препарат водой.
•Препарат высушивают на воздухе или фильтровальной бумагой.
•Препарат микроскопируют с помощью иммерсионной системы.

ОКРАСКА ПО МЕТОДУ БУРРИ-ГИНСА – для выявления капсул.
•Смешивают каплю культуры капсульных бактерий с каплей туши на конце предметного стекла.
•Затем готовят мазок как из капли крови.
•Мазок высушивают на воздухе.
•Фиксируют в пламени горелки.
•Окрашивают водным фуксином.
При этом способе окраски бактерии окрашиваются в красный цвет, а неокрашенные капсулы контрастно выделяются как светлый ободок на черно-коричневом фоне вокруг бактерий.
ОКРАСКА ПО МЕТОДУ НЕЙССЕРА – для выявления волютина.
•На фиксированный мазок наносят синьку Нейссера на 2-3 минуты.
•Не промывая водой, наносят раствор Люголя на 10-30 сек.
•Мазок промывают водой.
•Докрашивают раствором везувина в течение 1 минуты.
•Некоторые микроорганизмы содержат много зерен волютина (например дрожжеподобные грибы).
•Зерна волютина представляют собой соединения, имеющие, в отличие от цитоплазмы, щелочную реакцию, и поэтому окрашиваются в темно-синий цвет.
•Цитоплазма клетки, обладающая кислой реакцией, воспринимает щелочной краситель везувин и окрашивается в желтый цвет.
ОКРАСКА ПО МЕТОДУ ОЖЕШКИ – для выявления спор.
•На нефиксированный мазок наносят 0,5% раствор соляной кислоты и подогревают на пламени горелки в течение 2 минут до появления паров.
•Препарат промывают водой, высушивают и фиксируют.
•Докрашивают по методуЦиля-Нильсена.
•Споры бактерий после данной окраски приобретают красный цвет, а тело бактерий – синий.
ОКРАСКА ПО ГРАМУ
•На фиксированный мазок кладут полоску фильтровальной бумаги, пропитанную раствором генцианвиолета.
•Через 2 минуты полоску удаляют.
•Не промывая препарат водой, наносят раствор Люголя на 1 минуту. Затем раствор Люголя сливают.
•Препарат обесцвечивают спиртом 20-30 секунд.
•Препарат промывают водой.
•Окрашивают водным фуксином 2-3 минуты.
•Препарат промывают водой.
•Высушивают на воздухе или фильтровальной бумагой.
Грамположительные бактерии (Гр+) прочно фиксируют комплекс генцианвиолета и йода, не подвергаются обесцвечиванию этанолом и поэтому не воспринимают дополнительный краситель фуксин, оставаясь окрашенными в фиолетовый цвет.
У грамотрицательных бактерий (Гр+) этот комплекс легко вымывается из клетки этанолом, и они при дополнительном нанесении фуксина окрашиваются в красный цвет.
 15. МЕТОДЫ МИКРОСКОПИИ (ЛЮМИНЕСЦЕНТНАЯ, ТЕМНОПОЛЬНАЯ, ФАЗОВО-КОНТРАСТНАЯ,
15. МЕТОДЫ МИКРОСКОПИИ (ЛЮМИНЕСЦЕНТНАЯ, ТЕМНОПОЛЬНАЯ, ФАЗОВО-КОНТРАСТНАЯ,
ЭЛЕКТРОННАЯ).
ЛЮМИНЕСЦЕНТНАЯ МИКРОСКОПИЯ - основана на способности ряда веществ биологического происхождения или некоторых красителей светиться под действием падающего на них света.
•Микроорганизмы, содержащие хлорофилл, витамин В12, алкалоиды, некоторые антибиотики, обладают первичной люминесценцией.
•Клетки микроорганизмов, в которых люминесценция слабо выражена или отсутствует, обрабатывают специальными красителями - флуорохромами (акридиновый оранжевый, примулин, родамин и др.) в виде сильно разбавленных водных растворов: 1:500 -1:100000. Такие растворы слабо токсичны, что дает возможность изучать неповрежденную клетку.
ТЕМНОПОЛЬНАЯ МИКРОСКОПИЯ - основана на освещении объекта косыми лучами света (эффект Тиндаля). При таком освещении лучи не попадают в объектив, поэтому поле зрения выглядит темным.
•Если в исследуемом препарате содержатся клетки микроорганизмов, то косые лучи отражаются от их поверхности, отклоняются от своего первоначального направления и попадают в объектив. На интенсивно черном фоне видны сияющие объекты. Такое освещение препарата достигается использованием специального темнопольного конденсора, которым заменяют обычный конденсор светлопольного микроскопа.
•При микроскопировании в темном поле можно увидеть объекты, величина которых измеряется сотыми долями микрометра, что находится за пределами разрешающей способности обычного светлопольного микроскопа. Однако наблюдение за объектами в темном поле позволяет исследовать только контуры клеток и не дает возможности рассмотреть их внутреннюю структуру.
ФАЗОВО-КОНТРАСТНАЯ МИКРОСКОПИЯ - с ее помощью можно наблюдать живые объекты, которые имеют коэффициенты преломления, близкие к коэффициентам преломления среды.
•Прозрачные объекты видны более четко, чем в проходящем свете обычного светлопольного микроскопа. При отсутствии специального микроскопа обычный световой может быть оснащен специальным фазово-контрастным устройством, которое переводит фазовые изменения световых волн, проходящих через объект в амплитудные. В результате этого живые прозрачные объекты становятся контрастными и видными в поле зрения.
•С помощью фазово-контрастной микроскопии изучают форму, размеры, взаимное расположение клеток, их подвижность, размножение, прорастание спор микроорганизмов основанную на превращении невидимых фазовых изменений световых волн, вносимых объектом, в амплитудные, различимые глазом.
ЭЛЕКТРОННАЯ МИКРОСКОПИЯ. Позволяет наблюдать объекты, размеры которых лежат за пределами разрешающей способности светового микроскопа (0,2 мкм). Электронный микроскоп применяется для изучения вирусов, тонкого строения различных микроорганизмов, макромолекулярных структур и других субмикроскопических объектов.
•Обычный просвечивающий электронный микроскоп похож на световой, за тем исключением, что объект облучается не световым потоком, а пучком электронов, генерируемым специальным электронным прожектором.
•Полученное изображение проецируется на люминесцентный экран с помощью системы линз. Увеличение просвечивающего электронного
микроскопа может достигать миллиона, однако, для атомно-силовых микроскопов и это не предел.
 16. РОСТ И РАЗМНОЖЕНИЕ БАКТЕРИЙ.ФАЗЫ РАЗМНОЖЕНИЯ.
16. РОСТ И РАЗМНОЖЕНИЕ БАКТЕРИЙ.ФАЗЫ РАЗМНОЖЕНИЯ.
РОСТ БАКТЕРИЙ – формирование структурно-функциональных компонентов клетки и увеличение бактериальной клетки
РАЗМНОЖЕНИЕ – самовоспроизведение, приводящее к увеличению количества бактериальных клеток в популяции.
РАЗМНОЖЕНИЕ БАКТЕРИЙ
Бактерии размножаются путем бинарного деления пополам, реже путем почкования.
Гр+ бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки
Гр- делятся путем перетяжки, в результате образования гантелевидных фигур, из которых образуются две одинаковые клетки.
Делению клеток предшествует репликация бактериальной хромосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной нитью), приводящая к удвоению молекул ДНК бактериального ядра — нуклеоида.
Репликация ДНК происходит в 3этапа:
1)инициация,
2)элонгация (рост цепи)
3)терминация
РАЗМНОЖЕНИЕ БАКТЕРИЙ В ЖИДКОЙ ПИТАТЕЛЬНОЙ СРЕДЕ
•Размножаясь в питательной среде, бактерии потребляют питательные элементы, что приводит в дальнейшу истощению питательной среды и прекращению роста бактерий. Культивирование бактерий в такой системе называют периодическим культивированием, а культуру — периодической.
•Если же условия культивирования поддерживаются путем непрерывной подачи свежей питательной среды и оттока такого же объема культуральной жидкости, то такое культивирование называется непрерывным, а культура — непрерывной.
При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост культуры.
 Рост периодической культуры бактерий, ВЫРАЩИВАЕМЫХ НА ЖИДКОЙ ПИТАТЕЛЬНОЙ СРЕДЕ, ПОДРАЗДЕЛЯЮТ НА НЕСКОЛЬКО ФАЗ:
Рост периодической культуры бактерий, ВЫРАЩИВАЕМЫХ НА ЖИДКОЙ ПИТАТЕЛЬНОЙ СРЕДЕ, ПОДРАЗДЕЛЯЮТ НА НЕСКОЛЬКО ФАЗ:
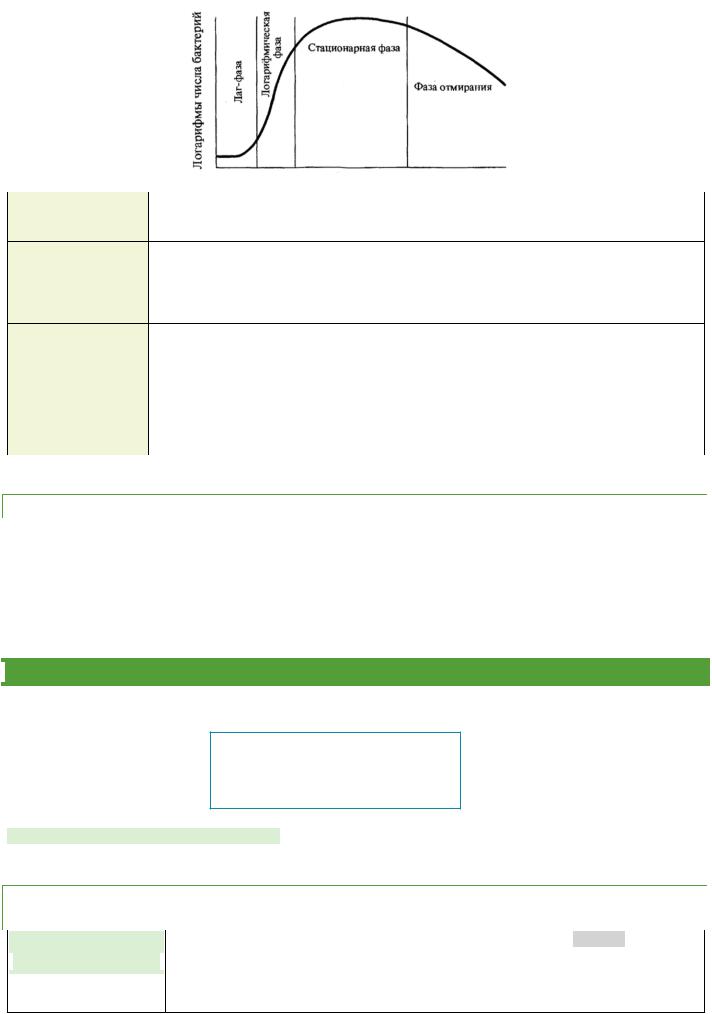
1. |
ЛАГ-ФАЗА |
период между посевом бактерий и началом размножения. Продолжительность лаг-фазы в |
|
|
среднем 4—5 ч. Бактерии при этом увеличиваются в размерах и готовятся к делению; |
|
|
нарастает количество нуклеиновых кислот, белка и других компонентов. |
2. |
ФАЗА |
период интенсивного деления бактерий. Продолжительность ее около 5— 6 ч. При |
ЛОГАРИФМИЧЕСКОГО |
оптимальных условиях роста бактерии могут делиться каждые 20—40 мин. Во время этой |
|
РОСТА |
фазы бактерии наиболее ранимы, что объясняется высокой чувствительностью компонентов |
|
|
|
метаболизма интенсивно растущей клетки к ингибиторам синтеза белка, нуклеиновых кислот |
|
|
и др. |
3. |
ФАЗА |
количество жизнеспособных клеток остается без изменений, составляя максимальный |
СТАЦИОНАРНОГО |
уровень (М-концентрация). Ее продолжительность выражается в часах и колеблется в |
|
РОСТА |
зависимости от вида бактерий, их особенностей и культивирования. |
|
4. |
ФАЗА ГИБЕЛИ |
отмирание бактерий в условиях истощения источников питательной среды и накопления в |
БАКТЕРИЙ |
ней продуктов метаболизма бактерий. Продолжительность ее колеблется от 10 ч до |
|
|
|
нескольких недель. Интенсивность роста и размножения бактерий зависит от многих фак- |
|
|
торов, в том числе оптимального состава питательной среды, окислительно- |
|
|
восстановительного потенциала, рН, температуры и др. |
РАЗМНОЖЕНИЕ БАКТЕРИЙ НА ПЛОТНОЙ ПИТАТЕЛЬНОЙ СРЕДЕ
Бактерии, растущие на плотных питательных средах, образуют изолированные колонии округлой формы с ровными или неровными краями (S- и R-формы), различной консистенции и цвета, зависящего от пигмента бактерий.
•Пигменты, растворимые в воде, диффундируют в питательную среду и окрашивают её.
•Другая группа пигментов нерастворима в воде, но растворима в органических растворителях.
•существуют пигменты, не растворимые ни в воде, ни в органических соединениях.
 17. СПОСОБЫ ПОЛУЧЕНИЯ ЭНЕРГИИ БАКТЕРИЯМИ (ДЫХАНИЕ, БРОЖЕНИЕ).
17. СПОСОБЫ ПОЛУЧЕНИЯ ЭНЕРГИИ БАКТЕРИЯМИ (ДЫХАНИЕ, БРОЖЕНИЕ).
К энергодающим процессам относят дыхание и брожение. Суммарные уравнения выглядят следующим образом:
С6Н12О6 +6О2=6СО2+6Н2О+674 ккал,
С6Н12О6 → 2СН3СН2ОН+2СО2+27 ккал.
ДЫХАНИЕ (БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ) - сложный процесс окисления различных органических соединений и восстановления некоторых минеральных соединений (нитратов и сульфатов).
ПО ТИПУ ДЫХАНИЯ МИКРООРГАНИЗМЫ РАЗДЕЛЯЮТ НА 4 ОСНОВНЫЕ ГРУППЫ: ОБЛИГАТНЫЕ АЭРОБЫ, ОБЛИГАТНЫЕ И ФАКУЛЬТАТИВНЫЕАНАЭРОБЫ И МИКРОАЭРОФИЛЛЫ.
ОБЛИГАТНЫЕ  (СТРОГИЕ) АЭРОБЫ
(СТРОГИЕ) АЭРОБЫ 
• растут при свободном доступе кислорода воздуха |
ПРИМЕР: |
(реакции окисления осуществляются у них при участии |
уксуснокислые |
молекулярного кислорода с высвобождением большого |
бактерии, |
количества энергии) |
возбудитель |
|
туберкулеза, |
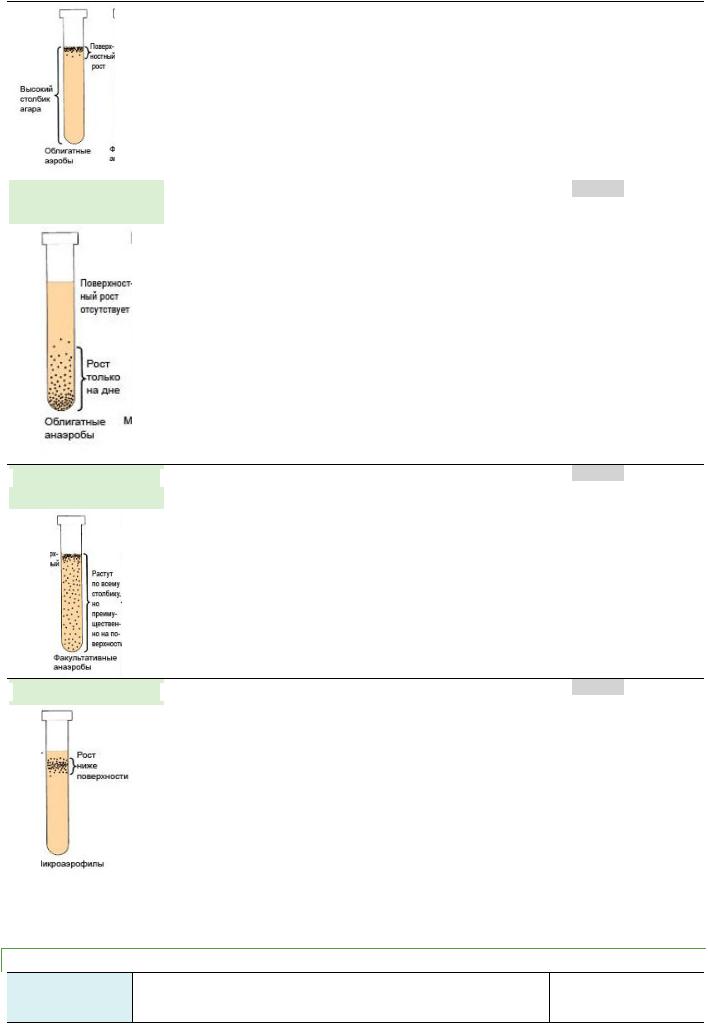
ОБЛИГАТНЫЕ
АНАЭРОБЫ
 ФАКУЛЬТАТИВНЫЕ
ФАКУЛЬТАТИВНЫЕ  АНАЭРОБЫ
АНАЭРОБЫ
 МИКРОАЭРОФИЛЛЫ
МИКРОАЭРОФИЛЛЫ
•Они имеют ферменты, обеспечивающие передачу водорода от донора электронов (субстрата) конечному акцептору - кислороду воздуха.
•Размножаются при наличии в атмосфере до 21% кислорода, на питательных средах растут на верхних слоях.
•способны к размножению только в атмосфере, свободной от кислорода. У этих м/о конечным акцептором водорода является субстрат (азотсодержащие вещества, углеводы и др.).
•растут на дне пробирки под значительным слоем питательной среды.
•Дыхание у анаэробов происходит путем ферментации субстрата с образованием небольшого количества энергии.
•Наличие свободного кислорода для облигатных анаэробов является губительным. Это связано с тем, что в присутствии кислорода конечным продуктом окисления органических соединений оказывается перекись водорода. Поскольку анаэробы не обладают способностью продуцировать фермент каталазу, расщепляющую перекись водорода, то она накапливается и оказывает токсическое действие на бактерии.
•развиваются как при доступе кислорода, так и в его отсутствии.
•имеют набор ферментов, обеспечивающих аэробный и анаэробный тип биологического окисления (дыхания).
•Развиваются по всей толщине питательной среды.
•нуждаются в значительно меньшем количестве кислорода, чем аэробы.
•Они развиваются при концентрации кислорода в окружающей среде не более 10%, т.е. у них преобладает аэробный тип дыхания.
ВИДЫ АНАЭРОБНОГО ДЫХАНИЯ
пигментные
гнилостные бактерии, многие плесени и др. микроорганизмы).
ПРИМЕР: маслянокислые и пропионовокислые бактерии, гнилостные клостридии, бифидобактерии и др.
ПРИМЕР:
молочнокислые
бактерии,
стафилококки, бактерии группы кишечной палочки, гнилостные бактерии рода Proteus.
ПРИМЕР:
актиномицеты,
лептоспиры,
возбудители
бруцеллеза,
некоторые плесневые грибы
НИТРАТНОЕ – |
- восстановления нитрата до молекул азота. |
у растений, грибов, |
диссимиляционная |
1) ассимиляция нитрата NО3- → NH3. |
бактерий |
нитратредукция |
В форме аммиака азот встраивается в органические молекулы. |
|