

Особенности восстановления половых клеток в ходе постнатального синтеа.
По способности к восстановлению половых клеток в ходе постнатального онтоенеза животные делятся на 3 группы:
Позвоночные, членистоногие и круглые черви, у которых единственным источником половых клеток являются гоноциты, которые образуются в ходе эмбриогенеза на ранних этапах развития и обособляются от соматических клеток и подвергаются необратимой детерминации, т.е. разрушение половых желёз у данных представителей в постнатальном развитии ведёт к бесплодию.
Ресничные черви, у которых запас половых клеток пополняется в течении всего постнатального развития, а клетки типа гоноцитов в ходе эмбриогенеза вообще не образуются. У них обнаружены малодифференцированные клетки – необласты, которые и являются источником половых клеток.
Кишечнополостные и губки относятся к этой же группе. У них имеются особые интерстициальные клетки, отличающиеся от необластов способностью дифференцироваться не только в половые, но еще и в половые клетки. Интерстициальные клетки пролиферируют всю жизнь в зависимости от условий обитания, но проявляют сезонный ритм.
 Кольчатые
черви, у которых в ходе эмбрион развития
на самых ранних этапах развития
образуются гоноциты, но при разрушении
половых желёз данные представители
не становятся стерильными, т.к у них
есть малодифференцированные клетки
типа необластов, способных в определённых
условиях дифференцироваться в половые
клетки.
Кольчатые
черви, у которых в ходе эмбрион развития
на самых ранних этапах развития
образуются гоноциты, но при разрушении
половых желёз данные представители
не становятся стерильными, т.к у них
есть малодифференцированные клетки
типа необластов, способных в определённых
условиях дифференцироваться в половые
клетки.
Особенности миграции гоноцитов.
Зачастую закладываются вдали от половых желёз, поэтому в ходе своего развития они мигрируют к развивающимся гонадами.
Как гоноциты, так и резервные клетки типа необластов способны двигаться самостоятельно, но при этом установлено, что значительную часть пути данные клетки проходят пассивно. Интерстициальные клетки с током воды в гастральную полость, а гоноциты куриного зародыша, напр., с током крови по эмбриональным кровеносным сосудам.
У млекопитающих есть своя особенность миграции. Она происходит в основном по интерстициальному типу, т.е активным образом. Они самостоятельно мигрируют из стенки желточного желтка, пробираясь через мезенхиму внезародышевых структур и мезенхиму тела зародыша к закладке тела желёз.
Миграция обусловлена образованием на поверхности гоноцитов псевдоподий.
В основе миграции лежат постоянные адгезивные отношения с элементами внеклеточного матрикса.
Процесс миграции гоноцитов детерменирован химическим сигналом – телофероном – гликопротеидной природы, обладающий неспецифическими свойствами, выделяется клетками поверхностного эпителия зачатка гонад (целомический эпителий).
Г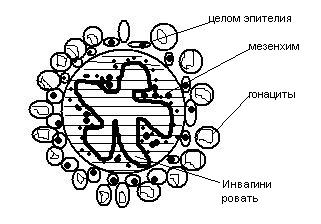 оноциты
могут мигрировать и с током крови (у
млекопитающих). Вблизи зачатка гонад
гоноциты путём диапедеза проникают
через стенку кров сосуда в мезенхиму
и дальнейший путь преодолевают
самостоятельно.
оноциты
могут мигрировать и с током крови (у
млекопитающих). Вблизи зачатка гонад
гоноциты путём диапедеза проникают
через стенку кров сосуда в мезенхиму
и дальнейший путь преодолевают
самостоятельно.
В ходе миграции половые клетки активно размножаются, их количество может увеличиваться в 50 раз.
На определённом этапе развития гоноциты достигают пол железы и устанавливают связь с целомическим эпителием. После этого целомический эпителий инвагинирует внутрь пол железы и увлекает за собой половые кл. В итоге зачатки половых клеток оказываются внутри половой железы и дальнейшее их развитие регулируется биологически активными веществами, которые выделяются соматическими клеток гонад.
Попав внутрь половой железы половые клетки продолжают активно делиться. В ходе развития женских половых клеток в период размножения овогонии не только делятся, но и подвергаются атрезии. В плоде до 7 млн овогоний, после рождения 1 млн овогоний, в дальнейшем 500тыс. При этом в ходе этапа размножения сперматогенеза сперматогонии активно делятся. У теплокровных деление идёт всегда, а у холоднокровных зависит от условий внешней среды.
Особенности питания яйцеклеток.
Женская половая клетка, прекратившая размножение – овоцит 1 порядка и вступает в период роста.
В периоде роста клетка должна получать большое количество питательных веществ, в ней усиливаются метаболические процессы, что обеспечивает её рост. У дрозофилы в периоде роста она увелич в 90к раз, у лягушек – 64к раз, у курицы – 200 раз. Столь интенсивный рост ЖПК в периоде роста потребовал выработки особых механизмов или типов питания.
Т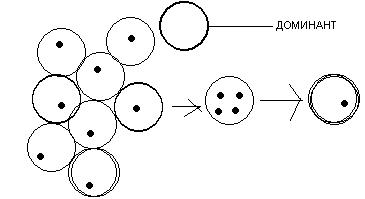 ипы
питания:
ипы
питания:
Фагоцитарный (губки): растущий овоцит ползает по межклеточному пространству, поглощая более мелкие клетки. Есть разновидность этого типа питания – двуступенчатый фагоцитарный тип пит – в этом случае овоцит присоединяет к себе клетку-носительницу, которая поглощает другие клетки или хоаноциты ближайшего … канала, а овоцит получает продукты расщепления
П
 ромежуточный
(кишечнополостные): овоциты развиваются
группами и в составе группы несколько
овоцитов становятся доминантными, т.е
они развиваются, поглощая сестринские
овоциты. После этого доминантные клетки
сливаются и образуется 1 многоядерный
овоцит, в котором в дальнейшем все ядра
кроме 1 разрушаются и образуется овоцит.
ромежуточный
(кишечнополостные): овоциты развиваются
группами и в составе группы несколько
овоцитов становятся доминантными, т.е
они развиваются, поглощая сестринские
овоциты. После этого доминантные клетки
сливаются и образуется 1 многоядерный
овоцит, в котором в дальнейшем все ядра
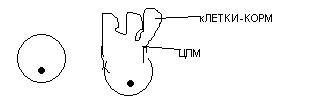
кроме 1 разрушаются и образуется овоцит.Нутриментарный (черви, членистоногие): обусловлен связью овоцита с клетками-кормилками. Причём кл-кормили имеют единый источник развития с овоцитом.
У дрозофиллы в ходе 4 кратного деления 1 овоцита образуются 16 клеток. 1 клетка становится овоцитом, а 15 – кормилки, которые связаны с овоцитом при помощи цитоплазматических мостиков, через которые к овоциту поступают питательные вещества.
П
 ри
помощи фолликулярных клеток
(млекопитающие): у насекомых данный
тип питания совмещается с нутриментарными,
а у млекопитающих он единственный.
ри
помощи фолликулярных клеток
(млекопитающие): у насекомых данный
тип питания совмещается с нутриментарными,
а у млекопитающих он единственный.
В отличии от клеток-кормилок фолликулярные клетки являются соматическими клетками и развиваются из целомического эпителия.
Также фолликулярные клетки не синтезируют питательный вещества, а только лишь их транспортируют из кровеносных сосудов к половым клеткам.
Фолликулярные клетки не образуют цитоплазматических связей с овоцитами. НО вместе с тем в ходе развития на поверхности фолликулярных клеток образуется отростки, которые инвагинируют в плазмолемму ЖПК, т.о. увеличивают площадь контакта, создавая переоцитарное пространство, куда выделяются питательные вещества.
Характеристика яйцеклеток.
О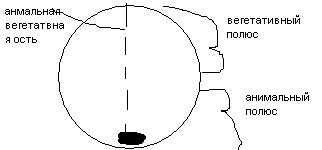 ткрыл
яйцеклетку Рапье де Грааф в 1672
– пузырьки на поверхности. Открытие
яйцеклеток. 1672 год- голландский анатом
и физиолог Рапье де Грааф.
ткрыл
яйцеклетку Рапье де Грааф в 1672
– пузырьки на поверхности. Открытие
яйцеклеток. 1672 год- голландский анатом
и физиолог Рапье де Грааф.
1826 год – Карл Бэр узнал, что пузырьки на поверхности — это вместилища, в которых развиваются ЖПК.
Женская половая клетка – большая клетка размером у лягушки 1,5-2 мм, у сельдевой акулы 20-25 см, у человека 120-140 мкм. Женская половая клетка образуется в ограниченном количестве у человека, в 1 половом акте - 1 женская половая клетка, а сперматозоидов 300-500 млн. Женская половая клетка зачастую неподвижна, не способна к самостоятельному передвижению, исключение – примитивные многоклеточные, у которых женская половая клетка способна к амебоидному движению.
Как правило, женская половая клетка имеет округлую/ овальную форму. Ядро располагается экстентрично. Ядро очень маленькое, расположено эксцентрично, округлое/ овальное с гаплоидным набором хромосом. Степень развития органоидов зависит от развития клетки. У взрослой развиты умеренно. У зрелой женской половой клетки нет клеточного центра. В цитоплазме есть кортикальные гранулы, располагающиеся по периферии под плазмолеммой. Данные гранулы окружены мембраной и содержат электронно плотное в-во и ферменты. Эти гранулы играют роль в образовании оболочки оплодотворения, т.е препятствуют полиспермии – проникновению более 1 сперматозоида. Больше всего этих гранул у яйцеклетки рыб.
Также в цитоплазме женской половой клетки есть пигментные гранулы (напр. у амфибий). Также есть углеводы, жиры и желток. Желток – фосфопротеиды, располагается в цитоплазме в виде кристаллов. Кол-во желтка – генетически детерминировано. Изменяя питание самки, не меняется кол-во желтка.
В женской половой клетке выделяют 2 полюса: анимальный, на котором располагается ядро, органоиды, пигмент гранулы, если есть; вегетативный полюс с основной массой желтка и трофическими включениями. Зрит ось, соединяющая полюса – анимально-вегетативная ось, которая является главной осью зародыша. При этом анимальный полюс соответствует головному отделу зародыша, а вегетативный- каудальному.
Для женской половой клетке характерны 3 вида оболочек:
Первичная оболочка – желточная. Образуется в ходе овогенеза за счёт уплотнения периферического и поверхностного слоёв цитоплазмы. Выполняет трофическую функцию.
Вторичные оболочки формируются за счёт клеток, которые окружают жпк. Это хитиновая оболочка у насекомых и блестящая у млекопитающих. Защитная функция.
Третичная оболочка формируется за счёт секрета желёз половых путей самки. Это белочная подскорлуповая и скорлуповая у птиц, кокон у червей, студенистая оболочка у лягушка. Защитная функция.
Классификация яйцеклеток по кол-ву и хар-ру распределения желтка.
А
 лецитальные
яйцелктки
– безжелтковые. Характерны для
паразитирующих форм: плоские черви,
печен сосальщик, эхинококк.
лецитальные
яйцелктки
– безжелтковые. Характерны для
паразитирующих форм: плоские черви,
печен сосальщик, эхинококк.Маложелтковые яйцеклетки – олиголецитальные/ изолецитальные. В данных яйцеклетках желток распределён по цитоплазме равномерно. Делятся на:
- первично изолецитальные (ланцетник). В основе разделения лежит эволюционный фактор.
- вторично изолецитальные (млекопитающие и человек).
Многожелтковые – полилецитальные/телолецитальные. Неравномерное распределение желтка, он сконцентрирован на вегетативном полюсе.
-умеренно (рыбы, амфибии).
-резко телолец (птицы).
Ц
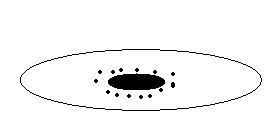 ентролецитальные
яйцеклетки
(насекомые). Вытянутая форма, ядро в
центре и в около ядерной зоне располагаются
гранулы желтка.
ентролецитальные
яйцеклетки
(насекомые). Вытянутая форма, ядро в
центре и в около ядерной зоне располагаются
гранулы желтка.
 СРАВНИТЕЛЬНАЯ
ЭМБРИОЛОГИЯ. ХАРАКТЕРИСТИКА ПОЛОВЫХ
КЛЕТОК.
СРАВНИТЕЛЬНАЯ
ЭМБРИОЛОГИЯ. ХАРАКТЕРИСТИКА ПОЛОВЫХ
КЛЕТОК.
Открытие сперматозоидов.
В 1877 Антони ван Левенгук описал в сперме "семенные пузырьки".
Термин сперматозоид ввел в начале ХХ века академик Петербургской Академии наук, немец по происхождению Карл Эрнст фон Бэр.
Сперматозоиды вырабатываются в мужском организме в большом количестве, по сравнению.с женскими половыми клетками. Так в эякуляте у собак до 60млн сперматозоидов; у лошадей до 10 млрд; у петуха 4 млрд; барана до 6 млрд; человек 350-500 млн сперматозоидов.
Сперматозоиды зачастую имеют жгутикообразную форму, что определяется их функцией. А именно, сперматозоиды Должны преодолеть большое расстояние до половой клетки и проникнуть в Женскую половую клетку.
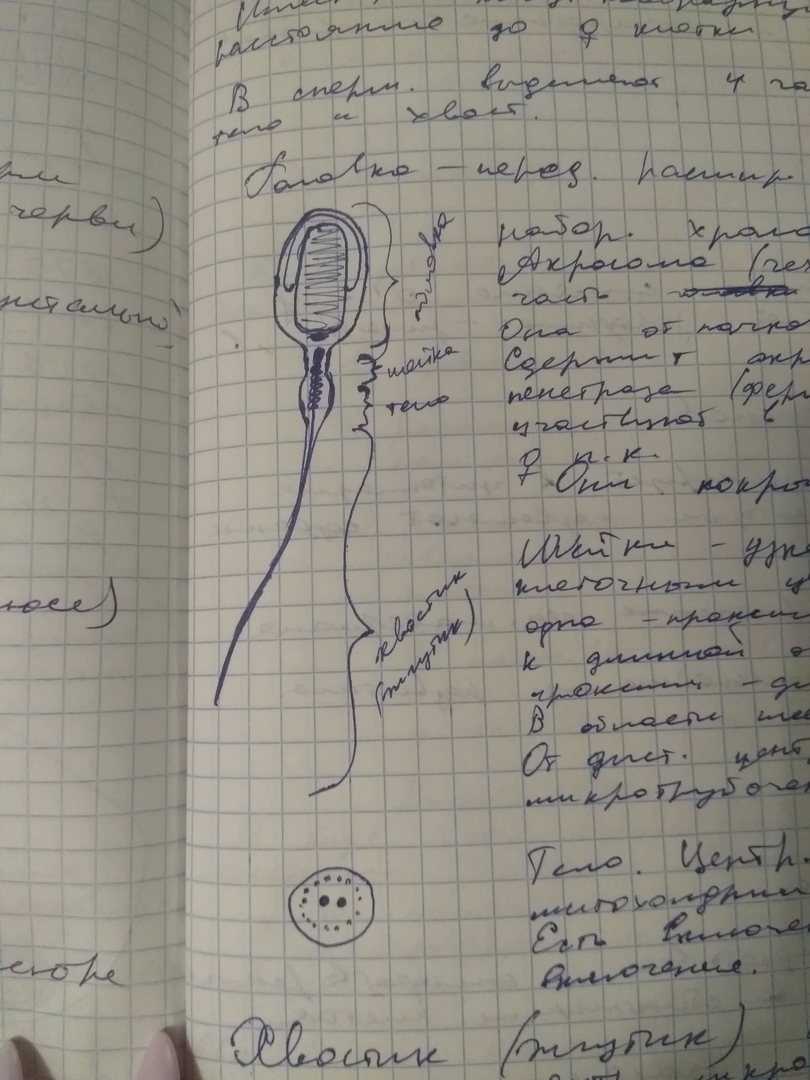
В сперматозоиде выделяют 4 части: головка, шейка, тело и хвост.
Головка- передняя расширенная часть сперматозоида, где располагается ядро. Может быть округлой, овальной формы в зависимости от вида сперматозоида. В ядре располагается гаплоидный набор хромосом.
Спереди или передний конец или Передняя часть ядра, ее 2/3 покрыта структурой-акросома,чехлик, или перфаратурий. Акросома является видоизмененной лизосомой. Акросома образуется и отпочковывается от транс-полюса Комплекса Гольджи и содержит целый комплекс ферментов: акрозин, трипсин, гиаларуронидаза, пенитраза. Данные ферменты называются спермализины. Они участвуют в разрушении и размягчении оболочек Женской Половой Клетки. С поверхности ядро акросома покрывается плазмолеммой, формируя переднюю часть это головка.
Шейка сперматозоида представляет собой узкую связующую часть, в которой располагается клеточный центр, состоящей из двух центриолей.
Одна центриоль проксимальная, плотно прилежит к ядерной оболочкой, и располагается перпендкулярно длинной оси ядра.
В торая центриоль клеточного центра отстоит от проксимальной центриоли и называется дистальная центриоль. Это узкая часть сеерматозоида называется шейкой. Роль этого отдела сперматозоида состоит в том, что при жгутикообразных формах строения сперматозоида в этом участке сперматозоид может делать повороты головки и изменять направление своего движения.
От дистальной центриоли растет осевая нить или аксонема состоящая из 10 пар микротрубочек.
В Теле спрематозоида центральное место занимает осевая нить, которую окружают митохондрии, компактно располагаясь в теле по спирали. Они располагаются не отдельно, а формируют митохондриальную нить. Кроме того В теле сперматозоида находится включения гликогена и макроэргические соединения(АТФ). Тело является энергетической станцией сперматозода. За счет работы митохондрий образуется энергия АТФ, которая необходима для перемещения и движения сперматозоида.
Каудальный отдел это хвостик или жгутик сперматозоида, который представляет собой видоизмененную ресничку.
Хвостик сперматозоида на поперечном сечении состоит из осевой нити или аксонемы, которая представлена 2 центральными микротрубочками и по переферии располагается 9 пар микротрубочек. Данная осевая нить покрывается в начальных отделах тонким слоем или ободком цитоплазмы, которая при удалении от тела сперматозоида истончается и с поверхности осевая нить покрыта плазмолеммой.
Таким образом, хвостик или жгутик сперматозоида состоит из осевой нити представленной 2 центральными микротрубочками, 9 парами переферических микротрубочек, тонкий ободок цитоплазмы покрывает аксонему и с поверхности располагается непосредственно плазмолемма. В состав аксонемы жгутика сперматозоида входят дэнеины, которые за счет энергии АТФ способны изменять свою конфигурацию и обеспечивать двигательную активность сперматозоида. Т.е. Различные движения маятникообразные, турбообразные движения хвостика сперматозоида, благодаря чему сперматозоид передвигается.
Термин Видоизмененная ресничка связан с тем, что в состав хвостика сперматозоида входят дополнительные сложноорганизованные цитоскелеты, которые начинаются уже с шейки сперматозоида и представлены сегментированными волокнами, несегментированными волокнами, волокнистыми влагалищами, которые образованы особенными структурами цитоскелета, состоящих из кератиноподобных белков, которые придают жгутику жесткость.
Сперматозоиды различных видов имеют ту или иную особенность.
Медуза, улитка морской еж, жук, рак, паук, рыб, жаба, чайка, петух, млекопитающих.
РАЗДЕЛ: СРАВНИТЕЛЬНАЯ ЭМБРИОЛОГИЯ.
ОПЛОДОТВОРЕНИЯ: ПОНЯТИЕ, ОБЩАЯ ХАРАКТЕРИСТИКА, ОСОБЕННОСТИ У РАЗЛИЧНЫХ ПРЕДСТАВИТЕЛЕЙ ЖИВОТНОГО МИРА.
Весь период эмриогенеза состоит из периодов:
1)Оплодотворение
2)Дробление
3)Имплантация
4)Гаструляция
5)Образование осевых и внезародышевых органов
6)Гистогенез
7)Органогенез
8)Системогенез
Оплодотворение.
Биологический смысл оплодотворения состоит в слиянии МУЖСКОЙ и ЖЕНСКОЙ ПОЛОВЫХ КЛЕТОК с образованием одноклеточного зародыша-зиготы, в которой восстанавливается диплоидным набор хромосом характерный для представителей определенного вида.
Оплодотворение выполняет 2 Функции:
Половая функция- обеспечивает передачу генов от родителей к потомкам.
2) Репродуктивная функция- в ходе оплодотворения в женской половой клетке инициирует в цитоплазме яйцеклетки процессы. реакций, которые позволяют продолжить развитие и создание нового организма.
Роль сперматозоида в ходе оплодотворения:
Специфическая функция. Внесение в яйцеклетку половинного генетического материала отца.
Активация женской половой клетки к дальнейшему развитию.
Процесс оплодотворения включает 2 стадии:
Осеменение.
Оплодотворение.
Биологический смысл осеменения: подготовка Мужских и Женских Половых Клеток к оплодотворению. Осеменение
Включает процессы направленные на сближение и активацию половых клеток. Осеменение включает в себя процессы связанные с сближением половых клеток и с их активацией. В ходе осеменения происходит сближение и активация как мужской так и женской половой клеткой.
Признаки осеменения:
Непременным условием осеменения, является участие большого количества сперматозоидов.
Это необходимо для того, чтобы вблизи Женской Половой Клетки сформиловалось большое количество ферментов, необходимого для разрушения и размягчения оболочек яйцеклетки. Участие в осеменении большого количества сперматозоидов обусловлено тем, что для размягчения и разрушения оболочек яйцеклетки необходимо большое количество ферментов, их концентрация которая может создать необходимое количество сперматозоидов. При этом большая концентрация сперматозоидов обусловлено еще тем, что в ходе сближения сперматозоидов с яйцеклеткой большая часть сперматозоидов гибнет. Избыточное количество тоже нарушит процессы оплодотворения. Фруктоза которая образуется в семенных пузырьках, образуется только для определенного числа сперматозоидов.
2) Видонеспецифичность.т.е. Активировать женскую половую клетку к дальнейшему развитию можно сперматозоидами другого вида. Но несмотря на то, что этот процесс видонеспецифический, половые клетки непрошедшие осеменения независимо от искусственного или естественного оплодотворения не могут приступить к оплодотворению.
3) Обязательный процесс. Осеменение обязательный процесс.
Выделяют 2 вида осеменения: наружное и внутреннее осеменение.
При Наружнем осеменении Половые клетки выделяются во внешнюю среду(представители: водные животные(рыбы), пиявки, саламандры).
Внутренее осеменение когда мужские половые клетки вводятся в половые пути самки. Представитель: млекопитающие.
В ходе осеменения половые клетки проходят 2 фазы взаимодействия:(осеменение состоит из 2 фаз):
1)Дистантное взаимодействие- это взаимодействие которое осуществляется на некотором расстоянии до соприкосновения гамет.
2)Контактное осеменение это то осеменение которое происходит при соприкосновении, при контакте, Женской Половой Клетки и Мужской Половых клеток.
Дистантное взаимодействие.
Смысл дистанстного взаимодействие это сближения Мужской и Женской Половых Клеток.
Факторы, способствующие дистанстному взаимодействию:
Подвижность половых клеток ( преимущественно сперматозоидов).
Разноименность мембранных электрических зарядов.
Хемотаксис. Движение спрематозоидов по градиенту концентрации химических веществ вырабатываемых Женской Половой Клеткой. Например: известно, что сперматозоидов папортникообразных по градиенту концентрации лимонной кислоты. Сперматозоиды кишечнополостных изменяют направление движения вблизи почек со зрелыми яйцеклетками.
Женские половые клетки движутся по направления химического вещества, ферментам, фертилизина, который вырабатываются лучитым венцом яйцеклетки или яйцелетки !! Порядка..
В ходе взаимодействий половые клетки выделяют химические вещества- ГАМОНЫ, которые воздействуют на гаметы своего или противоположного пола.(открыты Франком Лили в 1911г.). Гомоны вырабатываемые ЖПК-гиногомоны, МПК-андрогомоны.
Гинагомон ! это никомолекулярное вещество небелковой природы вырабатываемой оболочками яйцеклетки. Значение: Оно повышает двигательную активность сперматозоидов. и повышают вероятность их встречи с яйцеклеткой.
Гинагомон !! (фертилизин, изоагглютинин)- белковые вещества, которые вызывают склеивание сперматозоидов в реакциях с комплементарных андрогомоном !! молекулы которого встроены в поверхностную оболочку сперматозоида. Смысл: Предохраняют яйцеклетку от полиспермии, от проникновения в цитоплазму яйцеклетки более чем одного сперматозоида.
АНдрогомон !- вещество небелковой природы представляющее собой антагонистом гиногамона ! и подавляет двигательную активность сперматозоидов.
Андрогамон !! растворяет оболочки яйцеклетки его называют лизином и отождествляют с гиалуронидазой.
Контактные взаимодействия.
Под влиянием многочисленных сперматозоидов которые подошли и окружили яйцеклетку в ее цитоплазме происходят существенные физико-химические изменения: увеличивается проницаемость плазмолеммы, усиливается обмен ионами,повышается вязкость цитоплазмы и т.д.
Активация сперматозоидов при контакте с ЖПК может проходить в различных разновидностях. Например: у морских первичнохордовых животных активация сперматозоидов происходит по типу акромсомальной реакции, которая происходит в 2 вариантах:
При контакте сперматозоида с яйцеклеткой ,в плазмалемме сперматозоидов открываются натриевые каналы и в головке в цитоплазме сперматозоидов увеличивается концентрация ионов натрия и кальция, что стимулирует кальций-зависимый экзоцитоз, спермализинов из акросомы в переовариальное пространство. Под действием ионов кальция происходит кальций зависимый экзоцитоз, а именно акросома движется по направлению к плазмолемме головки, сливается с ней и происходит выделение ферментов лизосом из акросомы в переовариальное пространство.
И
 з
головки сперматозоида растет
акросомальная нить по направлению к
ЖПК, за счет перестройки актиновых
протофеламентов. Наружняя часть
акросомальной нити лизируется, и
ферменты выливаются в переовариальное
пространство.
з
головки сперматозоида растет
акросомальная нить по направлению к
ЖПК, за счет перестройки актиновых
протофеламентов. Наружняя часть
акросомальной нити лизируется, и
ферменты выливаются в переовариальное
пространство.
В дальнейшем из акросомальной нити растут акросомные трубочки, которые достигая плазмолеммы яйцеклетки сливаются с ней, формируют входные пути для дальнейшего проникновения ядра и клеточного центра сперматозоидов.
Акросомальную реакцию могут вызывать как контакт с твердой поверхностью половой клетки или оболочек ЖПК (у морского ежа).
У млекопитающих активация сперматозоидов проходит в 2 стадии:
Капацитация. Это приобретение способности к оплодотворению. Стимулируется химическими веществами белковой природы, Альбунинами, секретами половых путей самки. Под действием ферментов происходит химическая модификация пламолеммы головки и акросомы сперматозоида. В следствие чего плазмалемма становится проницаемой для ферментов.
Акросомальная реакция. Из акросомы в переовариальное пространство изливаются ферменты гиарунидаза, акрозин, приводящие к разрушению и размягчению оболочек ЖПК.
Собственно оплодотворения.
Б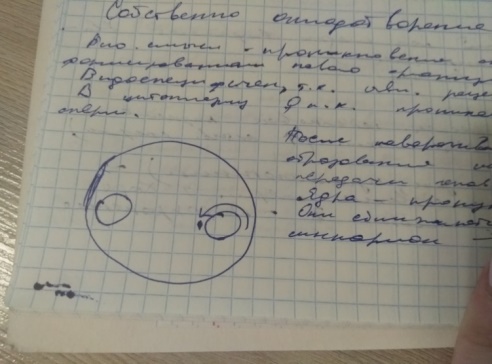 иологический
смысл
является проникновение сперматозоида
внутрь цитоплазмы яйцеклеток с
формированием генетически нового
одноклеточного организма. В отличии
от осеменения, оплодотворение является
видоспецифичный процесс, потому что
рецепторно опосредован. В цитоплазму
ЖПК проникает ядро и клеточный центр
сперматозоида.
иологический
смысл
является проникновение сперматозоида
внутрь цитоплазмы яйцеклеток с
формированием генетически нового
одноклеточного организма. В отличии
от осеменения, оплодотворение является
видоспецифичный процесс, потому что
рецепторно опосредован. В цитоплазму
ЖПК проникает ядро и клеточный центр
сперматозоида.
После проникновения сперматозоида: ядро сперматозоида поворачивается на 180 * клеточным центром вперед. Ядра на этой стадии называются -пронуклеосы. Постепенно ядра сближаются, сливаются и образуется -синкарио- одноклеточный организм зигота. И наступает 1 митотическое деление периода дробления.
В отличие от осеменения для оплодотворения необходим 1 сперматозоид- моноспермия или моноспермное оплодотворение характерно для млекопитающих.
В норме у некоторых представителей животного мира в цитоплазму ЖПК проникает 2 и более сперматозоида- полиспермиия или полиспермное оплодотворение. Характерно для членистоногие, рыбы. рептилии. птици, насекомые, пауки. Но с ядром ЖПК сливается ядро 1 сперматозоида и остальные погибают. Заканчивается моноспермией.
Полиспермное оплодотворение может встречатся у моноспермных, но в случае устарения, старения яйцеклеток. При проникновении это может привести к нарушению дальнейшего развития.
Механизмы блокады полиспермии:
Быстрая блокада полиспермии, связана с тем что при проникновении одного сперматозоида в цитоплазму яйцеклетки поступает небольшое количество ионов натрия что ненадолго именяет поверхностный заряд мембраны ЖПК и другие сперматозоиды проникнуть не могут.
Отсроченный механизм. Обусловлен участием кортикальных гранул.После проникновения первого сперматозоида из кальциевого депо выделяются ионы кальция движутся к плазмолеме ЖПК и по типу экзоцитоза выделяют свое содержимое формируя оболочки оплодотворения. Формирует плотную оболочку через которую не могут проникнуть сперматозоиды. Изменяет поверхностный мембранный заряд. Обусловлен действием ферментов кортикальных гранул на рецепторы ЖПК либо блокируют либо изменяют структуру рецепторов на поверхности яйцеклетки и сперматозоид не с чем взаимодействовать и они не проникают.
Женская половая клетка в ходе овогенеза претерпевает 2 стадии мейоза. И сперматозоиды, у различных представителей животного мира, в ходе оплодотворения проникают в ЖПК на различных этапах периода созревания. И исходя из этого выделяют 4 типа оплодотворения.
Типы оплодотворения
Первый тип (морской еж) - сперматозоид проникает в яйцеклетку лишь после прохождения обоих делений созревания.
Второй тип (позвоночные)- сперматозоид проникает в яйцеклетку на стадии метафазы !!-го деления созревания и необходим для завершения этого деления.
Третий тип (асцидии, моллюски) сперматозоид проникает в яйцеклетку на стадии метафазы ! деления созревания овогенеза.
Четвертый тип (Аскариды) сперматозоид проникает в яйцеклетку еще до прохождения обоих делений созревания.
 Овоплазматическая
сегрегация.
Овоплазматическая
сегрегация.
Ооплазматическая сегрегация – перемещение компонентов яйца после оплодотворения и формирование специфических участков («полей»), детерминирующим в дальнейшем развитие определенных частей зародыша.
Под ней понимают расслоение составных частиц цитоплазмы или ее интенсивное перемещение непосредственно после проникновения в цитоплазму ЖПК сперматозоида или воздействия партеногенетиеского фактора. У различных представителей животного мира имеет свои особенности.
У кишечнополостных сегрегация ограничивается расслоением цитоплазмы на 2 участка.
Это внешний ободок цитоплазмы- эктоплазма.
Внутренний ободок цитоплазмы- эндоплазма.
Эктоплазма бедна питательным материалом, но может содержать включения пигмента. Эндоплазма богата желтком. Такое простое расслоение цитоплазмы определяет дальнейшее развитие организма, а именно расположение нитей веретена делений в оплодотворенном яйце.
У асцидий сегргегационные
процессы хорошо выявляются благодаря
различной окраске составных частиц
цитоплазмы яйцеклетки.
асцидий сегргегационные
процессы хорошо выявляются благодаря
различной окраске составных частиц
цитоплазмы яйцеклетки.
В неоплодотворенном яйце у асцидий гранулы желтка равномерно располагаются по всей цитоплазме. После проникновения сперматозоида основная большая часть желтка смещается под экватор, с максимальной концентрацией небольшого участка его локализующей под экватором в виде структуры серповидной формы- желтый серп.
В результате этого цитоплазма анимального полюса характеризующийся бледно серым цветом, при этом над экватором располагается структура серповидной формы светло серой окраски, которая получает название светло серый серп.
Анимальный полюс в дальнейшем дает начало экодерме. вегетативный- энтодермы, желтый серп- мезодермы, светло серый -хорде.
Саггитальная ось соответствует главной оси зародыша.
Данные процессы определяют дальнейшие этапы развития зародыша: процесс дробления и гаструляции.
РАЗДЕЛ: СРАВНИТЕЛЬНАЯ ЭМБРИОЛОГИЯ.
ДРОБЛЕНИЕ: БИОЛОГИЧЕСКИЙ СМЫСЛ, ОБЩАЯ ХАРАКТЕРИСТИКА. ОСОБЕННОСТИ ДРОБЛЕНИЯ У РАЗНЫХ ПРЕДСТАВИТЕЛЕЙ ЖИВОТНОГО МИРА.
Биологический смысл дробления - образование многоклеточного организма - БЛАСТУЛЫ. Таким образом, из зиготы в ходе дробления образуется многоклеточный организм БЛАСТУЛА.
Основной процесс периода дробления является процесс митотическое деление клеток. При этом митоз ходе дробления имеет свою особенности: образующиеся в ходе деления дочерние клетки не растут, не достигают размеров материнской клетки, а с каждым последующим делением их размеры уменьшаются. В результате чего суммарная масса и объем бластулы равна объёму и массы зиготы.
Образующиеся в ходе дробления клетки получили название бластомеры (бласто-зародыш, мера-часть) что связано с тем, что данные клетки являются не дифференцированными или мало специализированными.
Особенности жизненного цикла бластомеров:
Бластомеры имеют сокращенную интерфазу, за счет отсутствия или сокращения G1 периода.
Удвоение ДНК начинается в телофазе предыдущего деления. Являются всегда тетраплоидными.
Уменьшение размеров бластомеры с каждым последующим делением происходит за счет уменьшения объемов цитоплазмы, при этом объем ядра не изменяется. Вследствие этого с каждым последующим делением происходит увеличение ядерно-цитоплазматического соотношения. Например: ядерно-цитоплазматическое соотношение у морского ежа в яйцеклетках 1:550, а в бластомерах 1:6.Считается, что процесс дробления заканчивается тогда когда в бластомерах создается определенное ядерно-цитоплазматического соотношения, необходимого для нормальной жизнедеятельности.
Второй биологический смысл дробления: восстановление ядерно-цитоплазматического соотношения. У половых клеток оно очень низкое, оно восстанавливается в периоде дробления.
 Процесс
дробления идет при помощи образования
особых специальных структур: борозды
дробления, перетяжка, инвагинация,
вдавления.
Процесс
дробления идет при помощи образования
особых специальных структур: борозды
дробления, перетяжка, инвагинация,
вдавления.
В дроблении участвуют 3 виды борозд дробления:
 Меридиональная.
Идет через анимально вегетативную
ось, которая соединяет анимальный и
вегетативный полюс.
Меридиональная.
Идет через анимально вегетативную
ось, которая соединяет анимальный и
вегетативный полюс.
Экваториальная (широтная). Идет перпендикулярно первой борозде, по экватору. Из-за особенностей содержания желтка может идти выше или ниже экватора.
Т
 ангенциальная.
Идет параллельно поверхности зародыша.
ангенциальная.
Идет параллельно поверхности зародыша.
Установлена прямая связь меду формированием нитей веретена деления и возникновением борозды дробления. При этом плоскость борозды дробления всегда совпадает с плоскостью метафазной пластинки. Считается, что положение будущей борозды дробления определяется в анафазе предыдущего деления. Есть предположения, что формированию борозды дробления способствуют инициирующие компоненты нитей веретена деления, которые обусловливают перестройку цитоскелета кортикального слоя цитоплазмы, вследствие чего на поверхности появляется инвагинация, которая распространятся по всей поверхности клетки и в глубину, что приводит к формированию перетяжки.
Гипотеза, высказанная Оскаром Гертвигом имела большое значение, для понимания дробления. Дробление определяется типом яйцеклетки. Чем больше содержания желтка в ЖПК, тем сложнее проходят борозды дробления. Большая концентрация желтка может привести к невозможности прохождения борозд дробления.
Исходя из этого, из зависимости высказанной Гертвигом, выделяют несколько типов дробления:
Полное или голобластическое дробление. Это когда все участки зиготы участвуют в дроблении, все дробятся.
Неполная или меробластическое дробление. Дробится определенный участок зиготы. Неполное дробление 2 вида:
2.1 -дискоидальное. Дробиться цитоплазма анимального полиса
2.2 -поверхностное. Дробится периферический участок цитоплазмы зиготы зародыша.
1*) Синхронное дробление. Борозды дробления с одинаковой скоростью проходят через все участки зиготы.
2*) Асинхронное дробление. Когда борозды дробления проходят через различные участки зиготы с неодинаковой скоростью или запаздывают на одном из полюсов зародыша.
1+)Равномерное это когда бластомеры, образующие в ходе дробления, имеют одинаковые размеры.
2+) Неравномерное. Образующиеся бластомеры отличаются по величине.
Виды полного дробления.
По характеру пространственного расположения бластомеров выделяют:
Радиальное. Характерно для Ланцетника, амфибии. При данном виде дробления образующиеся бластомеры располагаются друг, над другом создавая фигуру с латеральной симметрией, наподобие шара.
Спиральное. Характерно для Моллюски, кольчатые черви. При данном типе дробления вышележащие бластомеры смещены относительно ниже лежащих бластомеров под углом в 45*. Располагаются по спирали друг относительно друга.
Билатеральное. Характерно для Асцидии, нематоды. При данном типе дробления уже на ранних стадиях расположение бластомеров подчиняется закону билатеральной симметрии. т.е. бластомерам правой стороны соответствуют идентичные бластомеры левой стороны.
Х
 аотичное.
Характерно для некоторых медуз. При
данном типе дробления уже после 3
деления зиготы отсутствует строгая
закономерность в расположении
бластомеров. Возникает беспорядочность,
которое усиливается в ходе дальнейшего
дробления зиготы. При этом считается,
что данный тип дробления регулируется
внутренними механизмами, что приводит
к развитию и формированию организма
определенного вида.
аотичное.
Характерно для некоторых медуз. При
данном типе дробления уже после 3
деления зиготы отсутствует строгая
закономерность в расположении
бластомеров. Возникает беспорядочность,
которое усиливается в ходе дальнейшего
дробления зиготы. При этом считается,
что данный тип дробления регулируется
внутренними механизмами, что приводит
к развитию и формированию организма
определенного вида.
Таким образом, для дробления характерна пространственная организация бластомеров как относительно продольной оси зародыша, так и относительно друг друга. Эти проявления пространственной организации определяются количеством желтка в яйцеклетки, а также направлением и чередованием борозд дробления.
Зависимость между расположением желтка и прохождением борозд дробления были сформулированы в конце 19в. Оскаром Гертмером на основе правил предложенных ранее ботаником Саакс для растительных клеток. В своих правилах Саакс писал, что ядро в растительных клетках стремиться занять геометрический центр. А нити веретена деления ориентируются по наибольшему длинному поперечнику цитоплазмы клетки.
Правило О.Гертвига:
1) Клеточное ядро стремиться расположиться в центре свободной от желтка цитоплазмы.
2) Веретено клеточного деления стремиться расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы.
Данные правила действуют только на начальных этапах дробления. В дальнейшем вступает в действие другие механизмы регулирующие процесс дробления.
1-Взаимное давление бластомеров.
2-Изменения количество желтка в бластомерах.
 3-Различные
явления взаимовлияния частей
развивающегося эмбриона.
3-Различные
явления взаимовлияния частей
развивающегося эмбриона.