

Лекции Хомича Цитология
.pdfплода. Наприкінці розвитку плода жовтковий мішок втягується в кишкову трубку.
Розвиток амніона і серози пов’язаний з розвитком амніотичної складки. На другу добу інкубації або насиджування зародок занурюється в латебру, внаслідок чого позазародкові ектодерма і парієтальний листок мезодерми формують навколо нього кільцеподібну амніотичну складку, верхівка якої виступає над зародком. Складка з обох боків зародка розростається в напрямку його спинної ділянки. Її протилежні краї зливаються і утворюють дві оболонки: зовнішню – серозу і внутрішню – амніон. При цьому зовнішній шар стінки серози утворює ектодерма, внутрішній шар – парієтальний листок мезодерми, а в стінці амніона зовнішнім шаром є парієтальний листок мезодерми, внутрішнім – ектодерма. В ділянці заднього кінця зародка краї амніотичної складки не зростаються, що призводить до утворення серо-амніотичного каналу, через який білок надходить у порожнину амніона і проковтується плодом.
Амніон оточує тільки зародок. Між ними знаходиться амніотична порожнина, заповнена амніотичною рідиною, що продукується клітинами ектодерми стінки амніона. Амніотична рідина створює водне середовище, в якому розвивається зародок, захищає зародок від механічних пошкоджень і є його поживним матеріалом.
Сероза огортає зародок, позазародкові органи та білок. Вона прилягає до шкаралупи й бере участь у газообміні. Клітини ектодерми серози виділяють ферменти, що переводять мінеральні речовини шкаралупи в іони. Останні кровоносними судинами потрапляють в організм плода і використовуються для його росту. Між серозою з одного боку, амніоном і жовтковим мішком з другого боку знаходиться щілиноподібна порожнина – позазародковий целом.
Алантоїс починає формуватися на третю добу інкубації як випинання кишкової трубки. Його стінка, як і стінка жовткового мішка, утворена ентодермою та вісцеральним листком мезодерми. Алантоїс у вигляді ковбасоподібного мішка розростається в позазародковому целомі. Його мезодерма зростається з мезодермою серози, внаслідок чого утворюється серозо-алантоїс. У мезодермі серозо-алантоїса розвиваються кровоносні судини, через які до плода надходять Оксиген і мінеральні речовини. В порожнині алантоїса накопичуються продукти обміну речовин і сечова кислота. Протока алантоїса входить до складу пуповини, за допомогою якого плід сполучається із позазародковими органами.
Позазародкові органи функціонують тільки в період розвитку зародка (плода), а наприкінці його розвитку редукуються.
73
Стадії розвитку курчати. Ембріональний розвиток окремих видів птахів вивчено недостатньо. Нині найповніше досліджено розвиток зародка (плода) курей.
Н.П.Третьяков і М.Д.Попов на основі характеру живлення зародка розробили класифікацію розвитку курчати. Згідно з цією класифікацією, ембріональний розвиток курчати проходить п’ять стадій:
перша стадія – латебрального живлення – відбувається впродовж перших 30–36 год. інкубації. В цю стадію зародок живиться жовтком латебри. Джерелом енергії є глікоген. Кровообігу немає. Відбувається закладання серця і формування жовткового мішка;
друга стадія – жовткового живлення – триває з 30–36-ї години до 7–8-ї доби інкубації. Упродовж цієї стадії формуються позазародкові органи, в стінці жовткового мішка утворюються кровоносні судини. Розвиваються органи тіла зародка, серце починає скорочуватись, функціонує печінка, в організм надходить Оксиген;
третя стадія – живлення білком і дихання Оксигеном повітря –
триває з 7–8-ї доби по 18–19-ту добу інкубації. Для цієї стадії характерний інтенсивний розвиток алантоїса, який разом із серозою забезпечують зародок киснем та мінеральними речовинами. Продовжується також розвиток усіх органів тіла зародка. Наприкінці цієї стадії зародок стає сформованим плодом;
четверта стадія – використання зародком кисню повітря з по-
вітряної камери яйця – триває з 18–19-ї доби інкубації до наддзьобування. В цю стадію алантоїс редукується, плід відчуває потребу в кисні, продзьобує внутрішній шар підшкаралупової оболонки і починає дихати повітрям повітряної камери. У зв’язку з легеневим диханням починає функціонувати мале коло кровообігу;
п’ята стадія – вилуплення – триває з 20-ї по 21-у добу інкубації. Упродовж цієї стадії жовтковий мішок з вмістом впинається в порожнину кишечнику, інші плодові оболонки припиняють функціонувати і відмирають. Плід продзьобує шкаралупу і залишає яйце.
Існує і інша класифікація розвитку курчати. За нею розвиток курчати відбувається у чотири періоди: зародковий (1–7 доба інкубації), передплідний (8–12 доби інкубації), плідний (13–19 доби інкубації) і вилуплення (20–21 доби інкубації).
Ембріогенез плацентарних ссавців. Яйцеклітина плацентар-
них ссавців і відповідно зигота оліголецитальні, як у ланцетника. Однак ранні етапи їх ембріогенезу відрізняються. Так, гаструляція у плацентарних ссавців відбувається, як у птахів. Це пов’язано з тим, що на ранніх етапах ембріогенезу ссавців виявляються особливості їх становлення у філогенезі. Ссавці походять від рептилій, які, як і птахи,
74
мають полілецитальні яйцеклітини. Підтвердженням цього є яйцекладні ссавці, яйцеклітина яких також полілецитальна. Мала кількість жовтка в яйцеклітині плацентарних ссавців – вторинне явище, що є наслідком їх внутрішньоутробного розвитку і живлення за рахунок організму матері. Перехідною формою від яйцекладних до плацентарних ссавців є сумчасті ссавці. Їм властивий внутрішньоутробний розвиток, однак він дуже короткий і не супроводжується утворенням плаценти. У зв’язку з цим він закінчується на стадії, яка відповідає передплодовому періоду ссавців. Подальший розвиток передплодів сумчастих ссавців відбувається в шкірній сумці.
Запліднення у ссавців внутрішнє, відбувається у яйцепроводах. Дроблення зиготи починається як повне рівномірне, потім воно закінчується повним нерівномірним і відбувається асинхронно. Таке дроблення називають архаїчним. При дробленні кількість бластомерів зародка може бути парною і непарною. Під час дроблення утворюються два різновиди бластомерів – темні й світлі. Їх скупчення формують морулу. В морулі світлі бластомери розміщуються зверху, а темні – знизу. Світлі бластомери інтенсивно діляться шляхом мітозу і розміщуються на периферії морули навколо темних бластомерів. Між темними та світлими бластомерами утворюється порожнина, заповнена рідиною. З утворенням порожнини морула перетворюється на бластулу ссавців – бластоцисту. Її стінку утворює трофобласт. Він сформований світлими бластомерами і виконує трофічну та захисну функції. Всередині бластоцисти локально на стінці трофобласта розміщена група великих темних клітин, які утворюють зародковий вузлик – ембріобласт. З нього розвиваються тіло зародка та позазародкові органи.
Дроблення зиготи починається в яйцепроводах і закінчується в матці. У свині дроблення відбувається впродовж трьох діб, у вівці – чотирьох, у корови – восьми діб. Під час дроблення оболонки яйцеклітини зникають і живлення зародка здійснюється секретом епітеліоцитів яйцепроводів і матки, який всмоктується трофобластом і накопичується в порожнині бластоцисти.
Зародок у порожнині матки інтенсивно росте і збільшується в об’ємі. При цьому клітини ембріобласта розміщуються на стінці трофобласта і утворюють зародковий диск. На ньому, як i на зародковому диску птахів, виділяються зародковий щиток і позазародкова частина.
Трофобласт у жуйних набуває веретеноподібної форми, у свині й кобили – овальної. На його зовнішній поверхні утворюються первинні ворсинки, за допомогою яких він з’єднується зі слизовою оболонкою матки і всмоктує секрет маткових залоз – маткове молоко.
75
Процес з’єднання зародка зі слизовою оболонкою матки називають імплантацією. Імплантація у корів і овець здійснюється на 17-ту добу після запліднення, у кобили – на 40–70-ту, у свині – на 13-ту добу, у макаки – на 9-ту, у людини – на 6-ту добу.
Гаструляція, диференціація зародкових листків і осьових органів ссавців відбуваються так само, як і у птахів. Починається гаструляція ще до імплантації зародка в слизову оболонку матки. Одночасно з гаструляцією значні зміни спостерігаються з трофобластом. Частина трофобласта, що знаходиться над зародком, розсмоктується, а інша частина входить до складу позазародкової ектодерми. Внаслідок цього зародок деякий час перебуває в порожнині матки нічим не прикритий (критичний період розвитку).
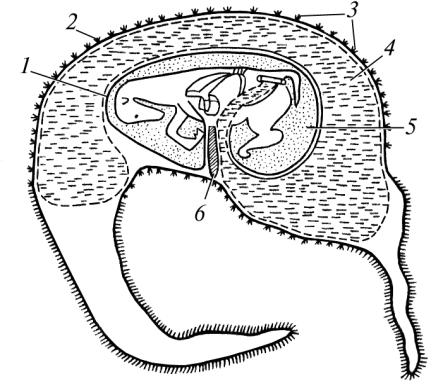
Позазародкові органи (плодові оболонки) ссавців утворюються із позазародкової частини зародкового диска. До них належать жовтковий мішок, амніон, алантоїс, хоріон і плацента. Джерела розвитку і функції жовткового мішка, амніона та алантоїса такі самі, як у птахів, а хоріон відповідає їх серозі (рис. 20).
Рис. 20. Плодові оболонки (схема):
1 — аланто-амніон; 2 — аланто-хоріон; 3 — ворсинки хоріона; 4 — порожнина алантоїса; 5 — порожнина амніона; 6 — жовтковий мішок
76
Жовтковий мішок, на відміну від такого птахів, не має жовтка, а містить білкову речовину. В зв’язку з цим його трофічна функція у жуйних і свиней незначна і він рано редукується. Так, у вівці жовтковий мішок існує з 13-ї по 30-ту добу розвитку, у корови – з 16-ї по 35ту, у свині – з 11-ї по 23-ю добу. У кобили жовтковий мішок у перші три місяці розвитку входить до складу плаценти, після чого також редукується.
Амніон утворюється у вівці на 15-ту добу розвитку, у корови – на 19-ту, у свині – на 13-ту добу і припиняє своє існування під час родів.
Алантоїс формується у вівці на 16-ту добу розвитку, у корови – на 20-ту, у свині – на 15-ту добу. Його мезодермальний шар зрощується з відповідним шаром хоріона й амніона, внаслідок чого утворюються аланто-хоріон та аланто-амніон. В останніх розвиваються кровоносні судини, які через пуповину сполучаються із судинами зародка.
Хоріон – це зовнішня плодова оболонка. На ньому формуються ворсинки (вторинні), в які вростають кровоносні судини. Ворсинки хоріона сполучаються зі слизовою оболонкою матки.
Плацента. Плацента утворюється наприкінці ембріогенезу ссавців й існує до кінця вагітності. Вона забезпечує нормальний розвиток зародка (плода) й виконує численні функції. Через плаценту в зародок (плід) надходять поживні речовини і Оксиген з материнського організму та виводяться продукти обміну. Вона захищає плід від дії механічних чинників, є органом його імунного захисту. В ній виробляються біологічно активні речовини, які регулюють нормальний перебіг вагітності та стимулюють розвиток молочної залози.
Плацента складається із зародкової (плодової) і материнської частин. Зародкову частину формує аланто-хоріон, у жуйних і амніохоріон, а материнська частина представлена слизовою оболонкою матки. У різних видів тварин плаценти відрізняються розміщенням ворсинок хоріона і характером їх сполучення зі слизовою оболонкою матки. Залежно від розміщення ворсинок хоріона розрізняють дифузну, котиледонову, кільцеподібну й дископодібну плаценти.
У дифузній плаценті ворсинки рівномірно розміщені по всій поверхні хоріона. Така плацента властива кобилі, свині, ослиці та верблюдиці.
Котиледонова плацента притаманна жуйним. Ворсинки хоріона цієї плаценти розміщені групами – котиледонами, які вступають у зв’язок з карункулами слизової оболонки матки.
77
У кільцеподібній плаценті ворсинки розміщені на хоріоні у вигляді смужки, яка оточує його. Ця плацента властива хижакам.
Дископодібну плаценту мають гризуни й примати. Ворсинки хоріона цієї плаценти займають площу, яка має вигляд диска.
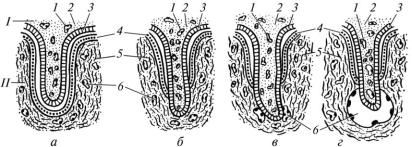
Залежно від характеру сполучення ворсинок хоріона зі слизовою оболонкою матки розрізняють епітеліохоріальну, десмохоріальну, ендотеліохоріальну та гемохоріальну плаценти (рис. 21).
В епітеліохоріальній плаценті ворсинки хоріона проникають у маткові залози і контактують з їх епітелієм. Така плацента властива кобилі, свині та ослиці. Для десмохоріальної плаценти характерно те, що ворсинки хоріона заглиблюються у товщу слизової оболонки, руйнуючи її епітелій. Ця плацента притаманна жуйним. Хижакам властива ендотеліохоріальна плацента. Ворсинки хоріона такої плаценти
Рис. 21. Типи плацент (схема):
а — епітеліохоріальна; б — десмохоріальна; в — ендотеліохоріальна; г — гемохоріальна; І — зародкова частина; ІІ — материнська частина; 1 — кровоносні судини ворсинки; 2 — сполучна тканина; 3 — епітелій хоріона; 4 — епітелій; 5 — сполучна тканина; 6 — кровоносні судини слизової оболонки матки
проникають у товщу слизової оболонки і контактують з ендотелієм кровоносних капілярів. При цьому вони руйнують епітелій і власну пластинку слизової оболонки матки. Гемохоріальна плацента притаманна гризунам і приматам. Ворсинки хоріона цієї плаценти заглиблюються в товщу слизової оболонки і проникають у розширені кровоносні капіляри, тобто вони контактують безпосередньо з кров’ю.
У плацентах усіх типів їх кровоносні судини й судини матері залишаються ізольованими. Між ними є гематоплацентарний бар’єр, непроникний для мікроорганізмів, токсинів і антитіл, що можуть міститися в крові матері.
Періоди внутрішньоутробного розвитку ссавців. Тривалість внутрішньоутробного розвитку свійських ссавців різна: у великої рогатої худоби вона становить 285–290 діб, у коней – 335–338, у овець – 150, у свиней – 114, у кролів – 30 діб. Внутрішньоутробний розвиток
78
ссавців поділяють на три періоди: зародковий (ембріональний), передплодовий і плодовий.
Зародковий період починається дробленням зиготи і закінчується утворенням плаценти, формуванням тканин і початком розвитку органів тіла зародка (органогенез). У великої рогатої худоби він триває в перші 35 діб вагітності, у овець – 25–30, у свиней – 23 доби, а у коней – перші три місяці вагітності.
Упередплодовий період закінчується формування плаценти, відбувається активний розвиток органів, їх систем і апаратів, у зв’язку
зчим збільшується маса передплода. Передплодовий період триває у великої рогатої худоби з 35-ї по 60-ту добу вагітності, у овець – з 30-ї по 45-ту, у свиней – з 23-ї по 38-му добу, у коней – у першій половині четвертого місяця вагітності.
Уплодовий період відбувається подальший розвиток органів, починається їх функціонування і збільшується маса плода. У великої рогатої худоби цей період триває з 60-ї доби вагітності до її закінчення, у овець – з 46-ї, у свиней – з 39-ї доби, а у коней – з середини четвертого місяця до закінчення вагітності.
Запитання для самоконтролю
1. Назвіть позазародкові органи птахів. 2. Джерела розвитку і значення жовткового мішка. 3. Як відбувається розвиток амніона і серози? .4. Які функції виконують амніон і сероза? 5. З чого розвивається алантоїс? 6. Яке значення алантоїса? 7. Стадії розвитку курчати. 8. Як відбувається дроблення зигот плацентарних ссавців? 9. Які особливості бластули плацентарних ссавців? 10. Особливості гаструляції плацентарних ссавців. 11. Що таке імплантація? 12. Назвіть позазародкові органи плацентарних ссавців. 13. Чим утворена плацента і які вона виконує функції? 14. Критерії класифікації плацент. 15. Періоди внутрішньоутробного розвитку ссавців.
79
ЗАГАЛЬНА ГІСТОЛОГІЯ
Лекція 10. Загальна характеристика тканин. Епітеліальна тканина
Предмет вивчення загальної гістології. Поняття про тканину. Типи тканин. Розвиток тканин (гістогенез). Закономірності гістогенезу. Регенерація тканин. Типи тканин. Загальна характеристика та мікро- і субмікроструктура епітеліальної тканини (епітелію). Класифікація епітеліальної тканини. Поверхневий епітелій і його різновиди. Залозистий епітелій. Залози, критерії їх класифікації. Секреція та її фази.
Загальна гістологія вивчає розвиток, будову та функції тканин організму. Тканина – це сформована в процесі історичного розвит-
ку система клітин та їх похідних, яка об’єднана спільністю по-
ходження, будови і функції. Тобто тканина складається із клітин та їх похідних (міжклітинної речовини).
Термін „тканина” вперше ввів у науку англійський вченийботанік Н.Грю у 1671 році. За допомогою мікроскопа він вивчав будову розплющених рослин і переплетіння їх волокон нагадувало йому тканину текстилю. У морфології тварин і людини цей термін запропонував використовувати французький морфолог К.Біша у 1801 році.
Процес розвитку тканин називають гістогенезом (грецьк. histos
– тканина, genesis – походження). Він починається після утворення зародкових листків і має низку закономірностей, основними з яких є диференціація, адаптація, інтеграція та проліферація. Клітини зародкових листків потрапляють у різні умови життєдіяльності. Так, клітини ектодерми розміщуються на поверхні тіла зародка, а клітини ентодерми – в його глибині. Між екто- і ентодермою лежать клітини мезодерми. У зв’язку зі зміною умов життєдіяльності спадкові (генетичні) можливості клітин реалізуються по-різному. Одні гени повністю реалізують свою інформацію, а діяльність інших блокується. Внаслідок цього шлях розвитку клітин змінюється, тобто настає їх диференціація, яка призводить до утворення нових популяцій клітин, що спеціалізовані для виконання певних функцій. Набуті в результаті диференціації нові морфофункціональні особливості клітин популяцій закріплюються в їхній спадковій інформації. В результаті цього здійснюється адаптація клітин до нових умов їх життєдіяльності. Створені нові популяції клітин інтегруються (об’єднуються) в тканини, з яких побу-
80
довані органи, їх системи та апарати. Одночасно кількість клітин в їх популяції зростає – проліферація.
Сформовані тканини в процесі життєдіяльності зазнають дії різноманітних чинників зовнішнього та внутрішнього середовища організму, що часто призводить до її руйнування. Частина клітин завершує свій життєвий цикл і також відмирає. Поновлення тканин називається регенерацією. Вона відбувається за рахунок стовбурових клітин. Оскільки у нервовій тканині таких клітин немає, вона не здатна до регенерації.
Існувало багато класифікацій тканин, але в більшості вони були примітивними, оскільки базувались не на мікроскопічних дослідженнях. У другій половині ХІХ століття німецькі вчені Ф.Лейдіг та Г.Келікер запропонували класифікацію тканин, якою користуються і нині. Підґрунтям її стали мікроскопічні і функціональні особливості тканин. Згідно з цією класифікацією, розрізняють чотири типи тканин: епітеліальну, сполучну, м’язову та нервову.
Епітеліальна тканина, або епітелій, є найбільш давньою тка-
ниною, що виникла на початку еволюції багатоклітинних організмів. Вона побудована з клітин – епітеліоцитів і майже не містить міжклітинної речовини. Епітеліоцити мають будову, яка властива соматичним клітинам. Особливістю їх є те, що вони мають органели спеціального призначення – тонофібрили. Епітеліоцити щільно прилягають один до одного і з`єднані різними типами контактів, серед яких обов`язково є десмосоми. Вони утворюють суцільний пласт, який знаходиться на базальній мембрані. Базальна мембрана є похідною епітеліоцитів і клітин пухкої волокнистої сполучної тканини, що розміщена під ними. Вона має товщину 1 мкм і утворена гомогенною основною речовиною та сіткою ніжних волокон. Базальна мембрана перешкоджає епітелію вростати в сполучну тканину і таким чином виконує бар`єрну функцію. Через неї відбувається живлення епітеліоцитів та виведення продуктів їх обміну. При цьому поживні речовини проникають до епітеліоцитів через базальну мембрану із крові судин сполучної тканини. Для епітеліоцитів характерна полярна диференціація. В них чітко виражені два полюси – апікальний і базальний. Апікальний полюс спрямований до зовнішнього середовища, а базальний знаходиться на базальній мембрані і з`єднаний з нею напівдесмосомами. Апікальний і базальний полюси епітеліоцитів мають і різні морфологічні ознаки. В базальному полюсі розміщені переважно ядро та окремі органели (мітохондрії, ендоплазматична сітка). Плазмолема епітеліоцитів цього полюса може утворювати глибокі впинання в цитоплазму. На апікальному полюсі багатьох різновидів епітеліо-
81
цитів є такі структури, як мікроворсинки, війки, щіточкова облямівка та ін. Полярна диференціація властива і пластам епітеліоцитів. Якщо пласт багатошаровий, то полярна диференціація притаманна лише епітеліоцитам найглибшого шару.
В епітеліальній тканині відсутні кровоносні і лімфатичні судини та є багато нервових закінчень. Вона має високу здатність до регенерації.
Епітеліальна тканина переважно займає пограничне положення в організмі. Вона вкриває шкіру, слизові і серозні оболонки, протоки залоз тощо (поверхневий епітелій) і утворює більшість залоз (залозистий епітелій). У зв`язку з цим вона виконує численні функції, які об’єднують у три групи: захисну, обміну речовин, секреторну.
Існує низка критеріїв класифікації епітеліальної тканини. Найпоширенішими серед них є морфологічний і генетичний (за походженням).
Морфологічний критерій ґрунтується на відношенні клітин епітеліальної тканини до базальної мембрани, що також зумовлено її функціями. За цими ознаками епітелій поділяють на простий (одношаровий) і багатошаровий. У простому епітелії всі епітеліоцити розміщені на базальній мембрані, а в багатошаровому – лише клітини найнижчого шару.
Простий епітелій поділяють на однорядний і багаторядний (псевдобагатошаровий). Однорядним називають епітелій, клітини якого лежать на базальній мембрані, вони мають однакову форму і розміри, а їхні ядра розміщені на одному рівні.
Псевдобагатошаровий епітелій характеризується тим, що всі його клітини розміщені на базальній мембрані, але вони мають різну форму і розміри, а їхні ядра знаходяться на різних рівнях і утворюють кілька рядів.
Багатошаровий епітелій поділяють на багатошаровий плоский зроговілий, багатошаровий плоский незроговілий та перехідний. Клітини багатошарового епітелію мають різну форму, однак визначення цього епітелію зумовлене формою і станом епітеліоцитів поверхневих шарів. У базальному шарі багатошарового епітелію знаходяться стовбурові клітини.
Генетичний критерій базується на походженні епітелію з різних зародкових листків. За цим критерієм розрізняють ектодермальний, ентодермальний і мезодермальний епітелій.
Ектодермальний (шкірний) епітелій розвивається з ектодерми.
Це багатошаровий і псевдобагатошаровий епітелій. До нього нале-
82