

Глава 2.2. Механизмы цитотоксичности
В основе токсического действия веществ лежит повреждение клеток, сопровождающееся их функциональными, либо структурно-функциональными изменения. Разнообразие формирующихся при этом эффектов со стороны целостного организма обусловлено сложностью организации клеток, многообразием клеточных форм, составляющих организм. Сформировавшиеся в процессе эволюции особенности структуры и функции отдельных клеточных типов, формирующих различные органы и ткани, настолько существенны, что чувствительность различных клеток к токсикантам может отличаться в тысячи раз. Тем не менее, живое объединено общностью фундаментальных свойств (см. выше), а это позволяет выделить и некоторые общие механизмы, лежащие в основе цитотоксического действия ксенобиотиков. К числу важнейших можно отнести следующие:
- нарушение энергетического обмена;
- нарушение гомеостаза внутриклеточного кальция;
- активация свободно-радикальных процессов в клетке;
- нарушение процессов синтеза белка и клеточного деления;
- повреждение клеточных мембран;
Необходимо отметить, что все эти механизмы тесно связаны друг с другом. Порой один из них является пусковым, но в дальнейшем особую значимость для судьбы поврежденной клетки приобретают другие. Очень часто два или несколько из упомянутых механизмов связаны между собой по типу «порочного круга». В этой связи их выделение носит искусственный характер и оправдывается только целями данного издания.
1. Нарушение процессов биоэнергетики
1.1. Системы энергообеспечения клетки
Если между двумя веществами осуществляется взаимодействие, например:
А + В « АВ,
то существует и некая константа равновесия, характеризующая силу этого взаимодействия:
К = [АВ]/([А] + [В]).
Если К = 1, то в равновесном состоянии компоненты реакции А (или В) и АВ находятся в смеси в одинаковых концентрациях. Если К > 1, то в смеси больше продукта реакции АВ, если К < 1, то, напротив, больше исходных ингредиентов А и В. Спонтанное взаимодействие А и В с образованием АВ происходит только тогда, когда К - велика.
Энергетика процесса взаимодействия веществ может быть охарактеризована следующим уравнением:
-RTlnKa = DG = DH - TDS, где
Ка -константа равновесия;
R - газовая константа (1,987 кал/град/моль);
Т - абсолютная температура;
G - свободная энергия;
Н - энтальпия (энергосодержание веществ);
S - энтропия (степень неупорядоченности системы).
Спонтанно протекают реакции, при которых свободная энергия системы в процессе преобразования уменьшается. Это может быть следствием двух явлений: уменьшение энтальпии или увеличения энтропии системы. В первом случае говорят об экзотермической реакции (DН > 0), так как она проходит с выделением энергии; во втором - об эндотермической. Если DG - положительно (свободная энергия системы возрастает), реакция не может идти спонтанно. Для того, чтобы такой химический процесс шел необходимо поступление энергии из окружающей среды, либо сопряжение его с другим, экзэргическим, процессом, например:
Х + АТФ « Х-АДФ + Ф DG (-)
Х-АДФ + У « ХУ + АДФ DG (-)
В этом случае эндэргический процесс:
Х + У « ХУ DG (+)
становится возможным за счет сопряженного экзэргического процесса:
АТФ « АДФ + Ф DG (-),
в ходе которого образуется промежуточный активный продукт Х-АДФ.
Жизнь клеток и макроорганизмов есть постоянный процесс синтеза сложных молекул (нуклеиновых кислот, белков, полисахаридов, липидов и т.д.), то есть структур с достаточно высокой энтальпией и низкой энтропией. Образование таких молекул означает увеличения свободной энергии системы. Следовательно синтетические процессы невозможны без одновременного протекания экзэргических реакций, обеспечивающих высвобождение энергии, поступающей из окружающей среды и запасенной в форме химических соединений. Основным видом таких реакций в организме является гидролитическое расщепление богатых энергией веществ, содержащих пирофосфатные связи (макроэрги), таких как: аденозинтрифосфорная кислота (АТФ), аденозиндифосфорная кислота (АДФ), гуанозинтрифосфорная кислота (ГТФ_, цитозинтрифосфорная кислота (ЦТФ), уридинтрифосфорная кислота (УТФ), ацилфосфаты и др. В конечном итоге энергетические возможности клетки определяются запасами макроэргов и, в первую очередь, АТФ.
АТФ постоянно синтезируется в ходе двух клеточных процессов: гликолиза и биологического окисления субстратов (клеточное дыхание). Оба находятся в тесном взаимодействии. Гликолиз проходит в цитоплазме клеток. Дыхание, основной путь образования макроэргов, осуществляется в митохондриях. В процессе дыхания, благодаря совместному действию большого числа ферментов, соответствующие субстраты, образующиеся при метаболизме белков, жиров, углеводов, поступающих с пищей, окисляются кислородом, также поступающим из окружающей среды, с образованием СО2, Н2О и энергии, выделяющейся в форме тепла и запасаемой в молекулах АТФ. Процессы, обеспечивающие поддержания определенного уровня АТФ в клетках организма, составляют основу и сущность их энергетического обмена.
Наиболее уязвимыми для действия токсикантов элементами биологической системы, обеспечивающей образование макроэргов в организме, являются: механизмы биологического окисления (ферменты цикла трикарбоновых кислот, ферменты дыхательной цепи), механизмы сопряжение биологического окисления и фосфорилирования (образования АТФ из АДФ и фосфата), механизмы доставки кислорода к клеткам кровью (рисунок 1).
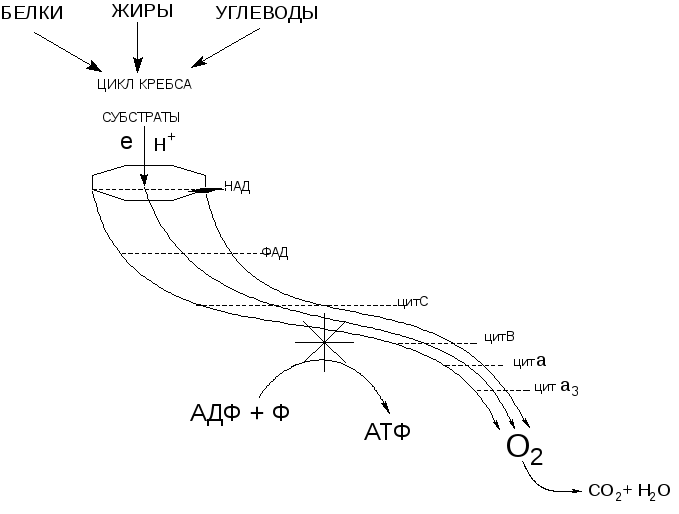
Рисунок 1. Гидродинамическая модель процесса клеточного дыхания. Энергия движения электронов и протонов (жидкость) по цепи дыхательных ферментов (желоб) обеспечивает работу механизма сопряжения (турбина) процесса фосфорилирования (кумуляция энергии)
Угнетение активности энзимов гликолиза редко приводит к существенному нарушению энергетического обмена, так как метаболизм других веществ (жиров, белков) может компенсировать этот вид нарушений.