

Основні іонні механізми виникнення ПС:
Наявність |
у |
клітинах |
|
Висока проникність |
|
Непроникність мембрани у |
трансмембранних градієнтів |
|
мембрани у стані |
|
стані спокою для |
||
концентрацій іонів К |
|
спокою для іонів калію |
|
внутрішньоклітинних білків- |
||
|
|
|
|
|
|
аніонів |
|
|
|
|
|
||
Дифузія іонів калію за градієнтом концентрації з клітини
Відносне зростання у позаклітинному |
|
Відносне зростання у цитоплазмі |
середовищі концентрації катіонів (+зарядів) |
|
концентрації аніонів (-зарядів) |
|
|
|
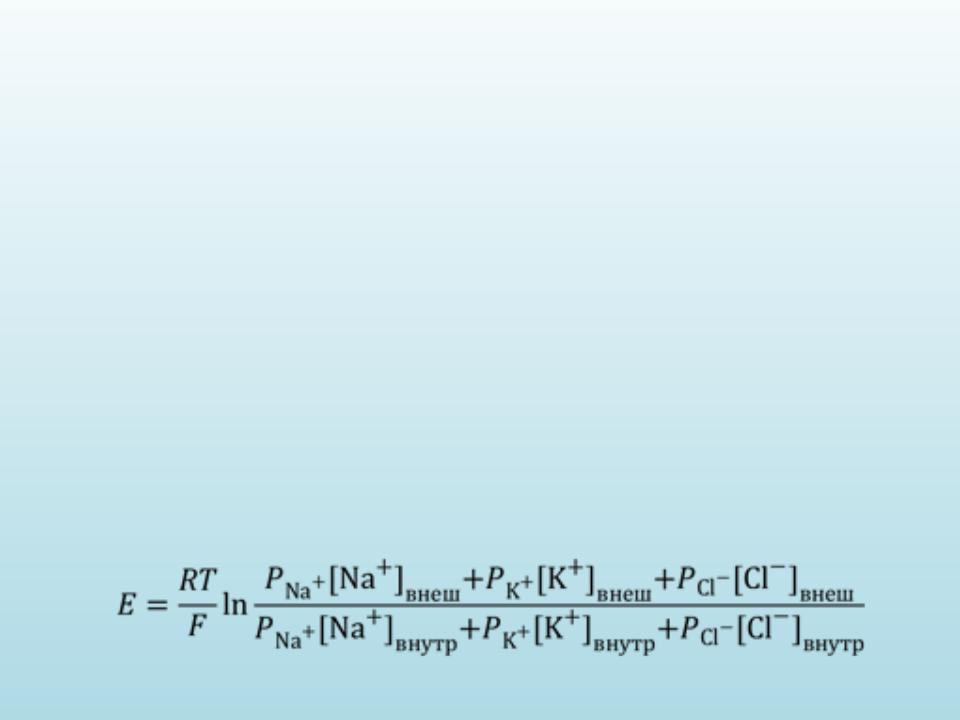
Більш точно підсумкова величина сумарного дифузійного потенциалу, який створюється витіканням деяких видів іонів, розраховується по формулі Гольдмана-Годжкіна-Катца.
В ній показано, що потенціал спокою залежить від трьох факторів: (1) полярності електричного заряду кожного іона; (2) проникності мембрани Р для кожного іона; (3) концентрації відповідних іоноі внутрі та ззовні мембрани. Для мембрани аксона кальмара у спокої відношення провідності РK : PNa :PCl = 1 : 0,04 : 0,45.

Відкриття електричних явищ у живих тканинах належить італійському вченому Гальвані (1759).
Мікроелектродний спосіб реєстрування біопотенціалів
(перший дослід Гальвані)
Скорочення литкового м’яза жаби при дії особливим пінцетом (Fe/ Cu) на сідничий нерв
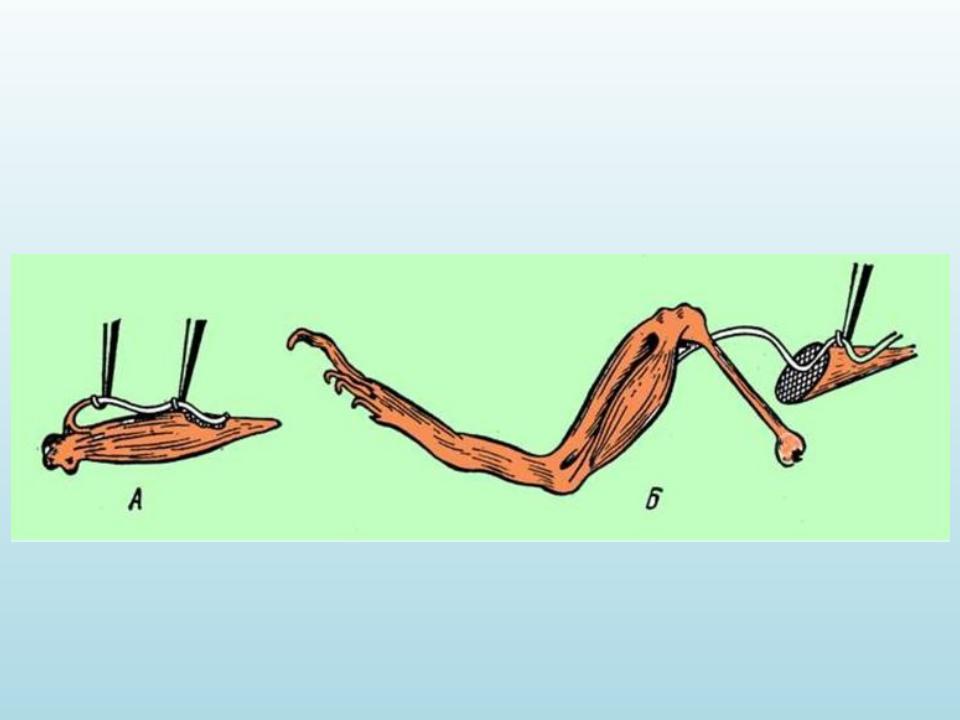
Другий дослід Гальвані:
Препарування сідничого нерва впродовж стегна лапки жаби і потім накидання його на м’язи гомілки за допомогою скляного крючка
У такий спосіб доведено, що джерелом електричного струму є самі тканини. Ушкоджена поверхня тканини має негативний заряд по відношенню до неушкодженої.

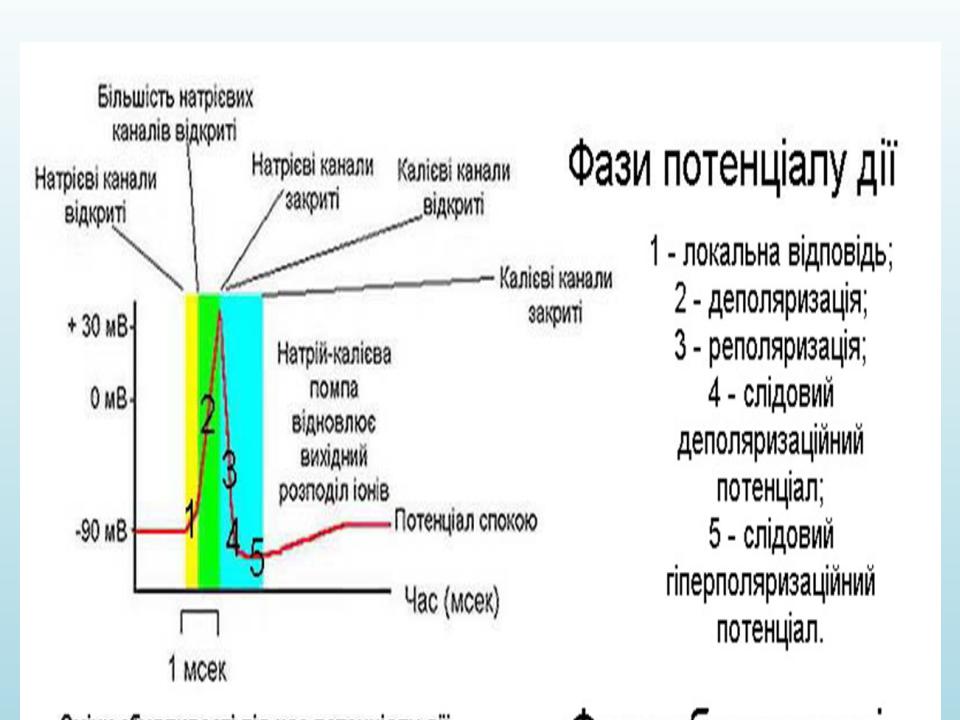
ПОТЕНЦІАЛ ДІЇ
Це короткочасна зміна різниці потенціалів між зовнішньою та внутрішньою поверхнями мембрани, що виникає в момент збудження.
В основі будь-якого потенціалу дії лежать такі явища:
1.Мембрана живої клітини поляризована — її внутрішня поверхня заряджена негативно по відношенню до зовнішньої.
2.Мембрана має вибіркову проникність — її проникність для різних частинок (атомів або молекул) залежить від їх розмірів, електричного заряду і хімічних властивостей.
3.Мембрана збудливої клітини здатна швидко змінювати
свою проникність, тобто для певного виду катіонів, викликаючи перехід позитивного заряду з зовнішньої сторони на внутрішню.
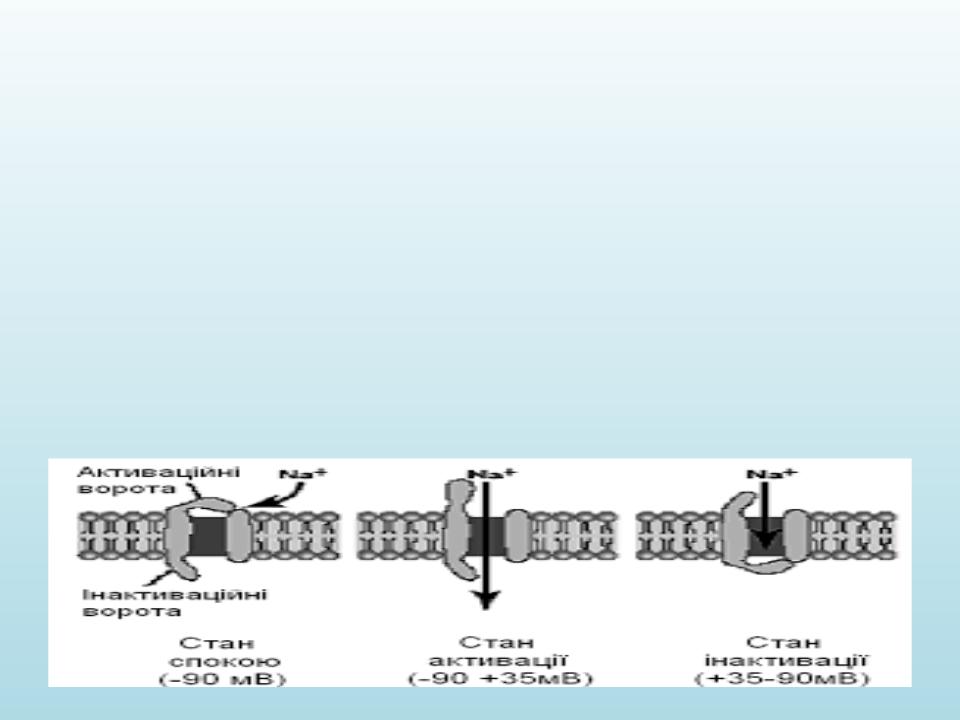
Молекулярні механізми виникнення потенціалу дії:
•Початкова фаза ПД формується вхідним натрієвим струмом, пізніше відкриваються калієві канали і вихідний K+-струм повертає потенціал мембрани до вихідного рівня. Початкову концентрацію іонів потім відновлює натрій-калієвий насос.
•По ходу ПД канали переходять зі стану в стан: у Na+-каналів
основних стани три — закритий, відкритий і інактивований, у K+ каналів два — закритий і відкритий.

Робота натрій-калієвого насосу:

Локальна відповідь:
Локальна відповідь – це не здатна до поширення деполяризація мембрани. В основному вона обумовлена переміщенням іонів Na+ в клітину. Рівень поляризації мембрани зменшується.
Особливості локальної відповіді:
1.Виникає при дії допорогових подразників.
2.Градуально залежить від сили деполяризуючого подразника.
3.Не здатна до розповсюдження.
Якщо сила подразника викличе таке підвищення проникності для іонів Na+ і мембрана зможе деполяризуватися не місцево, а вся, без будь-яких додаткових впливів, то виникає потеніцал дії, а такий подразник, що його викликав, називається пороговим, а сила подразника – порогом.
Величина мембранного потенціалу, з якої мембрана може продовжувати деполяризуватися автоматично називається
критичним рівнем деполяризації.
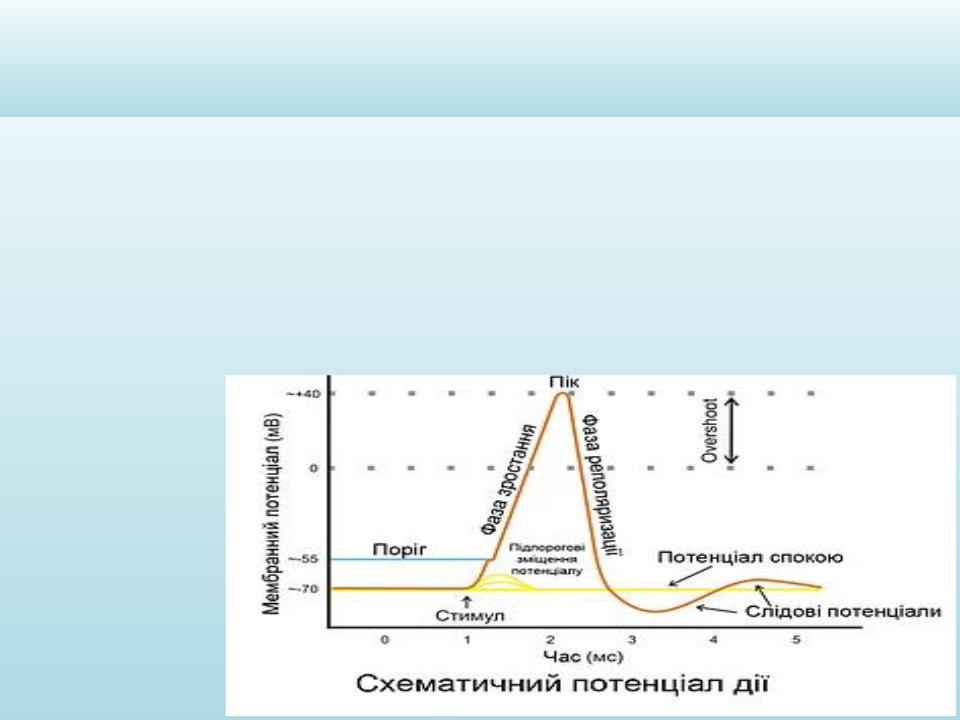
ДЕПОЛЯРИЗАЦІЯ:
Під дією подразника місцевий потенціал досяг критичного рівня
•При досягненні критичного рівня деполяризації (порога подразнення) відбувається Na – активація: відчиняються натрієві канали, Na надходить до клітини. Заряди на зовнішній та внутрішній поверхні мембрани спочатку вирівнюються, а потім відбувається зміна заряду мембрани.
•При досягненні певного значення ПД (120мВ) Na – канали закриваються, вхід Na зупиняється, але продовжується значний вихід К. Закінчується пік ПД.