

Строение микроскопических грибов
Клеточная стенка. Представляет собой многослойную оболочку из 9...10 слоев различной электронной плотности. Система микрофибрилл, встроенных в аморфный матрикс, формирует скелет клетки. Фибриллы в зависимости от видовой принадлежности могут состоять из целлюлозы, глюкона и хитина. Другие полисахариды, белки, пигменты, липиды служат цементирующими веществами, образующими химические связи с микрофибриллярной частью клеточной стенки. Наличие таких комплексов обеспечивает избирательную проницаемость для одних веществ и блокаду других.
Опорные микрофибриллы клеточной стенки и ее матрикс отличаются по механизму образования и биосинтезу. Образование фибрилл и матрикса происходит несинхронно, в первую очередь регенерируется фибриллярный остов стенки. Биосинтез этих двух частей клеточной стенки осуществляется с участием ферментов.
Процесс образования клеточной стенки происходит двумя способами: новый материал может либо внедряться в стенку поляризованно, либо равномерно накладываться по всей ее поверхности. В первом случае происходит образование цилиндрических клеток, во втором — сферических.
Клеточная стенка служит защитным приспособлением и предохраняет грибную клетку от воздействия различных факторов окружающей среды: осмотическим барьером, обусловливающим избирательную проницаемость для различных веществ. Она придает форму вегетативным клеткам гиф и органов размножения. На поверхности клеточной стенки и цитоплазматической мембраны локализованы ферменты, осуществляющие превращение не усвояемых клеткой (не растворимых в воде) полимеров.
В результате лизиса клеточная стенка грибов может разрушиться под воздействием ферментов, выделяемых другими клетками и образующихся в клетке самого гриба.
Основные компоненты клеточной стенки грибов — хитин, глюканы, белок и жиры. Азотистые и безазотистые полисахариды с жировыми веществами образуют растворимые и нерастворимые комплексы. Основу клеточной стенки составляют 4...6 моносахаров, соотношение которых у различных грибов незначительно варьирует. В состав полисахаридных фракций входят глюкозамин, манноза, глюкоза, ксилоза. Состав клеточной оболочки различных клеток одного и того же гриба неодинаковый.
Протопласт — содержимое клетки, заключенное в клеточную стенку: Имеет цитоплазматическую мембрану, эндоплазматический ретикулум, одно или несколько ядер с ядрышками, а также митохондрии, рибосомы с РНК, лизосомы, аппарат Гольджи, вакуоли, пластинчатый комплекс, секреторные гранулы, а также другие структуры и различные включения.
Цитоплазматическая мембрана. Тонкая трехслойная оболочка, располагается непосредственно под клеточной стенкой и отделяет ее от цитоплазмы. Цитоплазматическая мембрана обладает избирательной проницаемостью для веществ, входящих в клетку и выходящих из нее. Цитоплазматическая мембрана содержит до 40 % липидов и до 38 % белков. Различной формы инвагинации и ущемления цитоплазматической мембраны называются мезосомами.
Основное функциональное назначение цитоплазматической мембраны заключается в следующем: осуществление поступления в клетку различных веществ, ферментативная переработка и выделение продуктов метаболизма. Переработанные в цитоплазматической мембране вещества поступают в протопласт клетки и участвуют в обмене веществ.
Эндоплазматический ретикулум. Состоит из пузырьков, канальцев и вакуолей, служащих своеобразным депо питательных веществ.
Митохондрии. Многочисленные подвижные замкнутые образования эллипсовидной формы, с перегородками, покрытые одно- или двухслойной оболочкой. Предполагают, что митохондрии, благодаря собственной ДНК кольцевой структуры, способны к репродукции. Митохондрии окружены мембраной, на которой происходит локализация ферментов: пируватоксидазы, сукциндегидрогеназы, щелочной и кислой фосфатаз, пероксидазы. Митохондрии служат генераторами энергии в клетке. В зависимости от условий культивирования и физиологического состояния клетки форма митохондрий и их количество в клетке варьируют.
Рибосомы. Округлые зерна рибонуклеопротеидной природы, принимают участие в синтезе клеточных белков. Количество рибосом значительно отличается у различных видов грибов и зависит от внешних факторов, возраста культуры.
Аппарат Гольджи. Представлен группой пузырьков очень мелкого диаметра или параллельно лежащими дисковидными пластинками. Этот органоид располагается в клетке на участке, свободном от рибосом.
Лизосомы. Производные аппарата Гольджи, размешаются между клеточной оболочкой и цитоплазматической мембраной. Представляют собой зернистые образования, окруженные однослойной липопротеидной мембраной. Содержат фермент, гидролизирующий белок, и выполняют функцию зашиты клеток от неблагоприятного воздействия токсичных веществ экзо- и эндогенного происхождения.
Липосомы. Капельки жировых веществ, окруженные однослойными мембранами.
Ядро. Находится в центре или на полюсах клетки. В грибных клетках могут быть одиночные и множественные ядра. Они отвечают за наследственные функции. Форма ядер округлая или удлиненная. Каждое ядро окружено двухслойной пористой нуклеомембраной с ядрышком из плотных зерен и тонких фибрилл. Ядрышки содержат в составе хромосом ДНК. Через анастомозы ядра могут мигрировать из одной клетки в другую.
Включения. В грибных клетках многочисленные включения: волютин, гликоген, липиды, пигменты, миелоидные образования, соли органических кислот, аминокислоты. Гликоген ответствен за эндогенное дыхание, а волютин служит запасным питательным веществом, участвующим в энергетических процессах.
В процессе жизнедеятельности в клетках грибов накапливаются различные продукты метаболизма — антибиотики, ферменты, токсины, витамины.
Все многочисленные морфологические элементы микроскопических грибов подразделяют на две группы: мицелий и споры. Они бывают различной формы и размеров. Морфологическое различие спор и мицелия служит важным дифференциальным признаком при определении вида гриба.
Мицелий. Представляет собой узкую круглую трубку, диаметр которой варьирует у микромицетов от одного до нескольких микрон.
Ветвящиеся трубочки — гифы, составляющие мицелий, дифференцируют на более толстые слабо разветвленные и тонкие сильно ветвящиеся. Первые формируют мицелий, главным образом разевающийся на субстрате, вторые — в толще субстрата для поглощения из него питательных веществ. Такая дифференцировка особенно характерна для мицелия некоторых паразитных грибов, но встречается нередко и среди сапрофитных форм: у Rhizopus и некоторых других имеются особые столоны, неветвящиеся и обладающие энергичным ростом.
При обильном ветвлении гифы мицелия, соприкасаясь друг с другом, могут образовывать слияния между клетками, — анастомозы. При наличии большого их количества мицелий приобретает характерный сетчатый вид. Развитие анастомозов наблюдается у различных грибов с многоклеточным мицелием. Благодаря им возможно перемещение клеточного ядра из одной клетки в другую и переход от гаплоидного к диплоидному мицелию. Однако в большинстве случаев они осуществляют вегетативные функции и развиваются у многих форм при недостатке питания. Длина клеток мицелия колеблется от нескольких микрон до десятков и реже сотен микрон.
Мицелий окружен двухконтурной оболочкой, которая у молодых культур более нежная. В перегородках, делящих мицелий на отдельные клетки, имеются поры, через которые в процессе роста переливается цитоплазма, а с ней и питательные вещества. В клетках много различных включений: в старых цитоплазма становится зернистой из-за множества вакуолей. Молодой мицелий состоит из удлиненных прямоугольных клеток, старый — из коротких округлых или многогранных. Мицелий, имеющий перегородки, называется септированным. Однако у некоторых низших грибов мицелий состоит из гиф, лишенных поперечных перегородок, и представляет собой как бы одну, сильно разветвленную гигантскую клетку с многочисленными ядрами и называется несептированным мицелием.
Как же происходит развитие мицелия? Из споры выпячивается ростковая трубочка, которая удлиняется и затем отчленяется перегородкой от средней части, включающей спору. Ростовые трубочки затем еще удлиняются и получают новую перегородку, разделяясь на дистальную, или верхушечную, клетку и проксимальную, или внутреннюю. В дальнейшем верхушечная клетка удлиняется и вновь делится, отделяя вторую, более молодую по сравнению с первой, внутреннюю клетку. Так повторяется и дальше. В этом процессе внутренние клетки только вытягиваются, поперечное деление их происходит редко, но зато из них развиваются боковые ветви. На дистальном конце внутренней клетки образуется боковое выпячивание, принимающее цилиндрическую форму и отделяющееся затем перегородкой от производящей ее клетки. Новая клетка вырастает затем в боковую ветвь, растушую и ветвящуюся таким же образом, как и главная. Благодаря развитию ветвей на протяжении главной гифы они тем старше и сильнее развиты, чем ближе к основанию лежит то их отхождения — акропетальное ветвление.
Развитие несептированного мицелия происходит в общем таким же образом, но без образования поперечных перегородок. Рост происходит на кончиках гиф, где накапливается обильная протоплазма, заполняющая весь просвет, а в более задних частях происходит значительное развитие центральных вакуолей. В однородной среде, например на поверхности питательной желатины, гифы мицелия (как неклеточного, так и многоклеточного) разрастаются равномерно и радиально, так что мицелий имеет форму круга, нарастающего с краев. Центральная часть в нем самая старая, даже иногда отмершая, а периферическая — наиболее молодая.
При общем однообразии развития мицелия, который можно назвать типичным, в отдельных случаях наблюдается ряд специфических черт как макроскопического вида и общего характера роста, так и микроскопического строения. Макроскопический вид мицелия определяют прежде всего воздушные гифы. В одних случаях они формируются на самой поверхности субстрата и отчасти внутри его и тогда мицелий имеет вид плоского, прижатого к субстрату кружка; в других случаях, кроме того, развиваются более или менее обильные гифы, поднимающиеся в воздух и придающие мицелию некоторое сходство, например с куском ваты, возвышающимся над субстратом. Характер роста может быть различным у одного и того же гриба в зависимости от влажности, питания. Однако ряд форм грибов имеет специфические особенности, например образование пышного воздушного мицелия — разрушителя древесины.
Цвет мицелия чаше всего бывает снежно-белый, но с возрастом приобретает бурую окраску разных оттенков. Это связано с отложением пигмента в клеточных стенках и реже внутри самой клетки.
Различают мицелий истинный и псевдомицелий (отдельные клетки не связаны друг с другом и не имеют общей оболочки). Вместо истинного ветвления здесь наблюдается древовидное расположение клеток.
Для прикрепления к субстрату и извлечения из него питательных веществ в ходе эволюции у некоторых грибов сформировались специально предназначенные для этого органы: ризоиды и аппрессории, которые учитывают при идентификации грибов. Ризоиды — это корешкообразные, а аппрессории — короткие расширенные, иногда лопастеобразные выросты мицелия.
Грибы, паразитирующие на растениях, иногда формируют специальные ответвления мицелия — гаустории, которые, проникая непосредственно в клетки растений, обеспечивают питание грибов. Для гаусторий типично резкое изменение характера их роста по сравнению с ростом типичного мицелия, что объясняется воздействием протоплазмы живой клетки хозяина.
Склероции, тяжи, ризоморфы и хламидоспоры также являются видоизменениями мицелиального роста.
Склероции представляют собой септированные гифы грибов, образующие особые тела. При формировании склероциев оболочки гиф утолщаются и приобретают темную окраску. Сильно утолщена стенка гиф наружного слоя склероция, внутри же гифы более тонкостенные и обычно не окрашены. Склероции — это защитные приспособительные тела, которые позволяют грибу длительное время сохраняться в окружающей среде и обеспечивают его устойчивость к воздействию различных внешних факторов: температуры, солнечных лучей и др. Зрелые склероции содержат меньше влаги по сравнению с мицелием и много запасных веществ — липидов, гликогена.
Размеры склероциев колеблются от нескольких миллиметров до нескольких десятков сантиметров, а форма бывает самая разнообразная: сферическая, неправильная, в виде прямых или изогнутых рожков и др.
Структура клеток склероциев и механизм их образования различны, однако их формирование происходит путем увеличения ветвления мицелия и септирования гиф. Известны два способа образования склероциев: терминальный — на концах гиф; интеркалярный — в отдельных фрагментах главных гиф.
У многих грибов при развитии плодовых тел и некоторых вегетативных структур образуется ложная ткань — плектенхима (псевдопаренхима). В отличие от настоящей ткани паренхимы, возникающей в результате деления клеток в трех направлениях, плектенхима образуется путем сплетения и срастания. Если она состоит из клеток более или менее изодиаметрических, то ее называют параплектенхимой; если в ней заметно явное гифообразное строение (клетки удлиненной формы), то ее называют прозоплектенхимой.
Мицелиальные тяжи — вегетативная структура линейно агрегированных гиф. Диаметр мицелиальных тяжей зависит от количества гиф, которые концентрируются вокруг центральной основы.
В простейшем случае небольшое количество параллельно идущих гиф склеиваются друг с другом ослизненными наружными оболочками или вступают в более прочное соединение путем формирования многочисленных коротких анастомозов. В других случаях, когда тяжи массивны, их гифы получают определенную дифференцировку. Наружные элементы бывают более тонкими, образуя как бы кору вокруг центрального толстого ствола.
Ризоморфы — более сложные по агрегации гифы, которые отличаются у различных грибов интенсивностью роста центральной гифы, протяженностью боковых ветвлений, а также степенью дифференциации клеток гиф.
Наружные части у ризоморфы обычно темноокрашены и имеют определенное сходство с корнями высших растений. Они широко распространены у грибов с крупными плодовыми телами: у базидиальных, сумчатых и др.
Основное назначение мицелиальных тяжей и ризоморф состоит в обеспечении распространения грибов в субстрате и передвижении по гифам питательных веществ.
Хламидоспоры — это изменения мицелия в зрелых и старых культурах на концах или по его ходу. Основная функция хламидоспор не размножение, а сохранение вида. Форма их обычно круглая, овальная или слегка удлиненная, диаметр превышает диаметр мицелия. У некоторых грибов стенка двухконтурная, поверхность гладкая или шероховатая. Хламидоспоры могут возникать на концах мицелия, тогда они называются терминальными, по ходу мицелия — интерполярными (промежуточными).
В старых культурах часто наблюдают большие скопления хламидоспор причудливой формы, напоминающей четки или ожерелье. Молодые и зрелые хламидоспоры способны прорастать. Старые клетки дегенерируют.
Споры. С помощью спор грибы не только размножаются, но также и распространяются в окружающей среде. Этому способствует высокая устойчивость оболочек спор к воздействию агрессивных факторов. Споры подразделяют на эндоспоры, образующиеся внутри особых вместилищ — спорангиев (сумок), и экзоспоры, располагающиеся на мицелии.
У совершенных грибов споры подразделяют на ооспоры, зигоспоры, аскоспоры, базидиоспоры, эндоспоры, фиалоспоры, хламидоспоры. Споры несовершенных грибов в соответствии с размерами и происхождением также делят на несколько групп. К эндоспорам, образующимся внутри мицелия путем сегментации последнего, относят таллоспоры, включающие в себя артроспоры, хламидоспоры и бластоспоры. Кроме того, для несовершенных грибов характерно образование конидий, макроконидий, алейрий (микроконидий) и гемиспор, считающихся несовершенными конидиями.
Гемиспоры более прочно связаны с мицелием и представляют, собой один или два сегмента, отшнуровывающихся после поперечного деления мицелиальной нити. Форма их цилиндрическая, иногда округлая или многогранная, оболочка двухконтурная.
Морфология и строение плесневых грибов К плесневым грибам относят микроорганизмы, имеющие одноклеточный или многоклеточный мицелий, дифференцированные органы плодоношения, содержащие экзо- или эндоспоры. Они являются растительными гетеротрофными, хемоорганотрофными, аэробными организмами, лишенными хлорофилл. С растениями их связывает наличие клеточной стенки и вакуолей, заполненных клеточным соком, ток протоплазмы, отсутствие способности к движению. Однако по сравнению с растениями они слабо дифференцированы.
Вегетативное тело (мицелий) гриба состоит из длинных, разветвленных нитей или гифов, наполненных цитоплазмой. У ряда грибов мицелий образован гифами без поперечных перегородок (несептированный мицелий). Большинство грибов имеют гифы с поперечными перегородками, разделяющими их на участки (септированный мицелий).
Клеточная стенка большинства плесневых грибов содержит хитин или близкие к нему соединения. Под клеточной стенкой находится цитоплазма зернистой структуры в виде гранул рибосом. В цитоплазме грибов присутствуют митохондрии, в которых локализуются дыхательные ферменты, жировые включения и зерна волютина.
Клетки грибов обладают дифференцированным ядром, окруженным мембраной. Несептированный мицелий содержит несколько ядер.
Грибы размножаются, как правило, бесполым или половым способом. Способ размножения дает возможность определить систематическое положение того или иного гриба.
Б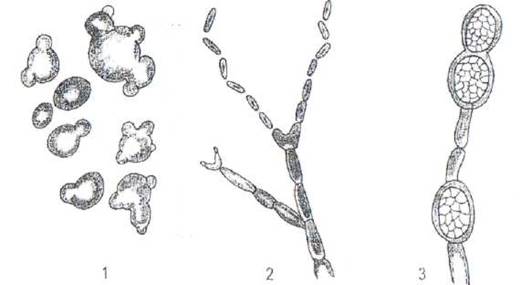 есполое
размножение происходит обычно при
помощи спор, а также путем почкования
или фрагментации (рис. 15), Наиболее широко
у грибов распространено спорообразование.
Споры грибов (конидиоспоры) отшнуровываются
от конидиеносцев или образуются
внутри спорангиев (спорангиоспоры). У
низших грибов спорангиослоры часто
движутся с помощью жгутиков. Движущиеся
спорангиоспоры носят название зооспоры.
есполое
размножение происходит обычно при
помощи спор, а также путем почкования
или фрагментации (рис. 15), Наиболее широко
у грибов распространено спорообразование.
Споры грибов (конидиоспоры) отшнуровываются
от конидиеносцев или образуются
внутри спорангиев (спорангиоспоры). У
низших грибов спорангиослоры часто
движутся с помощью жгутиков. Движущиеся
спорангиоспоры носят название зооспоры.
Рис. 15. Бесполое размножение грибов.
1- путем почкования (дрожжи); 2 - путем распада гиф на отдельные клетки - оидии или артроспоры; 3 - путем образования толстостенных хламидоспор (Fusarium).
Для дрожжей характерным видом бесполового размножения является почкование. При этом на материнской клетке образуется небольшая почка, в которую переходит одно ядро, после чего почка отшнуровывается.
Половое размножение у грибов напоминает размножение эукариотных клеток и включает слияние двух ядер.
Классификация грибов основана главным образом на способах размножения. Грибы подразделяют на пять классов, в которые входят низшие и высшие грибы. Низшие грибы (архимицеты и фикомицеты) объеденены в два класса, а высшие (аскомицеты, базидиомицеты и несовершенные грибы) - в три класса.
Архимицеты - наиболее примитивные грибы. У одних видов имеется зачаточный мицелий, представляющий собою вегетативное тело, состоящее из разветвленных переплетающихся нитей, называемых гифами. Тело других состоит из голого комочка цитоплазмы, который, превращаясь в спорангий со спорами, покрывается оболочкой. Большинство видов этого класса является внутриклеточными паразитами низших и высших растений. Некоторые из них поражают капустную рассаду («черная ножка»), клубни картофеля (рак).
Фикомицеты - грибы с одноклеточным несептированным, многоядерным и сильно разветвленным мицелием, который образует на питательной среде характерный войлочный налет. Размножаются фикомицеты половым и бесполым путем. Бесполое размножение осуществляется эндоспорами, заключенными в особых спорангиях. Половое размножение происходит в результате коньюгации двух половых клеток - гамет.
В класс фикомицетов входит род мукоспоровой (Mucor) или головчатой плесени (рис. 16). Тело ее состоит из нерасчлененного мицелия в виде сильно разветвленной клетки, от которой ответвляются плодоносящие воздушные гифы - спорангиеносцы с шарообразным расширением наверху - спорангиями. Спорангии заполняются эндоспорами, несущими функцию размножения. При недостатке питательных веществ мукоровая плесень может размножаться и половым путем. Она широко распространена в природе, часто обнаруживается на овощах, на влажных поверхностях, в навозе.
Рис. 16. Мукоровая плесень
А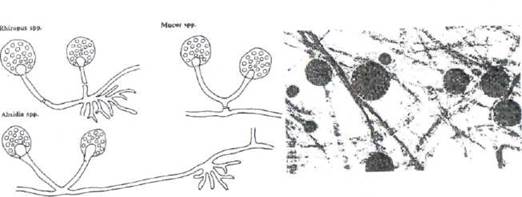 скомицеты,
или сумчатые грибы, представляют собой
самый обширный класс грибов. Они
характеризуются сильно разветвленным
многоклеточным мицелием. Размножение
у них обычно бесполое и осуществляется
при помощи конидий (экзоспор). При
половом процессе они размножаются
аскоспорами, развивающимися после
слияния ядер половых клеток (гамет) в
особых сумках - «асках».
скомицеты,
или сумчатые грибы, представляют собой
самый обширный класс грибов. Они
характеризуются сильно разветвленным
многоклеточным мицелием. Размножение
у них обычно бесполое и осуществляется
при помощи конидий (экзоспор). При
половом процессе они размножаются
аскоспорами, развивающимися после
слияния ядер половых клеток (гамет) в
особых сумках - «асках».
К классу аскомицетов принадлежат мицелиальные грибы. Из них чаще всего встречаются различные виды плесневых грибов аспергиллус (Aspergillus) и пенициллиум (Penicillium). Воздушные гифы этих грибов заканчиваются наверху утолщением, имеющим особые выросты - стеригмы. На концах стеригм образуются экзогенные вегетативные споры - конидии (рис 17).
Р ис.
17. Мицелиальные грибы
ис.
17. Мицелиальные грибы
Аспергиллы имеют многоклеточный (септированный) мицелий и одноклеточный конидиеносец. На верхнем конце конидиеносца веерообразно расположен ряд коротких стеригм, от которых цепочками отшнуровываются конидии (рис. 17.1).
При микроскопическом исследовании плодоносящая часть аспергиллов напоминает собой струйки воды, выливаемой из лейки. Это определило название - леечная плесень.
Мицелий и конидиеносец пенициллов - многоклеточные; плодоносящее тело имеет вид кисточки (рис. 17.2). В верхней части конидиеносец двукратно кистевидно разветвлен. На концах его образуются стеригмы, от которых отшнуровываются четкообразные ряды конидий.
Мицелий у пенициллов и аспергиллов не окрашен, а конидии могут окрашиваться в различные цвета, чаще всего в черный, сине-зеленый или серо-зеленый.
Аспергиллы и пенициллы широко распространены в природе. Типичным представителем аспергиллов является Aspergillus niger. который встречается на влажных предметах, в хлебе, варенье. Отдельные виды аспергиллов могут вызывать у человека аснергиллез дыхательных путей, глаз, уха и общее поражение организма.
Пенициллы обнаруживаются в кормах, молочных продуктах, на влажных предметах, в варенье. Некоторые виды этого гриба являются патогенными для человека. Они вызывают поражения кожи, ногтей, уха, верхних дыхательных путей, легких и других органов.
К классу Ascomycetes порядку Piectascales (первичносумчатые грибы) принадлежат дрожжи, которые представляют собой крупные одноклеточные неподвижные микроорганизмы (рис. 18).
О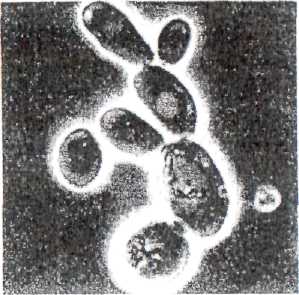 ни
образуют клетки овальной, шаровидной
и палочковидной формы, имеют
двуконтурную оболочку, митохондрии
и хорошо дифференцированное ядро.
Цитоплазма их гомогенна, иногда
тонкозернистого строения, имеет
включения (гликоген, волютин, жир) и
вакуоли, а также нитчатые тельца -
хондриосомы, которые принимают участие
в синтетических процессах, протекающих
в клетке. Дрожжи размножаются
почкованием, делением, спорообразованием,
а некоторые виды дрожжей - половым
путем. При почковании от материнской
клетки отделяются дочерние клетки,
превращающиеся в самостоятельные особи.
Отделение выростов-почек от материнских
клеток может происходить в любой стадии
развития.
ни
образуют клетки овальной, шаровидной
и палочковидной формы, имеют
двуконтурную оболочку, митохондрии
и хорошо дифференцированное ядро.
Цитоплазма их гомогенна, иногда
тонкозернистого строения, имеет
включения (гликоген, волютин, жир) и
вакуоли, а также нитчатые тельца -
хондриосомы, которые принимают участие
в синтетических процессах, протекающих
в клетке. Дрожжи размножаются
почкованием, делением, спорообразованием,
а некоторые виды дрожжей - половым
путем. При почковании от материнской
клетки отделяются дочерние клетки,
превращающиеся в самостоятельные особи.
Отделение выростов-почек от материнских
клеток может происходить в любой стадии
развития.
Дрожжи могут размножаться и спорообразованием. Споры образуются внутри клеток, оболочка которых выполняет роль сумки (аскуса). Это позволило отнести дрожжи к сумчатым грибам - аскомицетам. Споры у дрожжей округлой формы и покрыты гладкой оболочкой. В одном аскусе количество спор может достигать 2, 4, 6, 8, реже 12.
Многие виды и разновидности дрожжей обладают способностью сбраживать различные углеводы. Поэтому они широко используются в пивоваренной, винодельческой промышленности, хлебопечении.
К аспорогенным дрожжам относятся патогенные для человека виды из рода Candida, которые вызывают тяжелые заболевания - кандидозы. Они возникают вследствие подавления нормальной микрофлоры антибиотиками, применяемыми для лечения ряда инфекционных заболеваний и воспалительных процессов, а также при тяжелых заболеваниях, когда происходит снижение защитных сил организма.
Б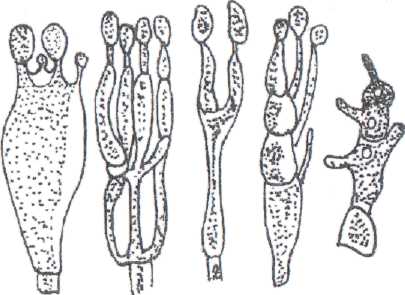 азидиомицеты -
грибы с многоклеточным мицелием (рис.
19). Они имеют дифференцированное ядро
и размножаются преимущественно половым
путем с помощью базидиоспор, которые
развиваются в специальных органах
размножения - базидиях. Базидии содержат
определенное количество спор, обычно
четыре. Большинство базидиомицетов
являются сапрофитами и живут на
перегнойной почве, на растительных
остатках, активно участвуя в разложении
органических соединений. Отдельные
виды являются паразитами сельскохозяйственных
растений, поражают древесину.
азидиомицеты -
грибы с многоклеточным мицелием (рис.
19). Они имеют дифференцированное ядро
и размножаются преимущественно половым
путем с помощью базидиоспор, которые
развиваются в специальных органах
размножения - базидиях. Базидии содержат
определенное количество спор, обычно
четыре. Большинство базидиомицетов
являются сапрофитами и живут на
перегнойной почве, на растительных
остатках, активно участвуя в разложении
органических соединений. Отдельные
виды являются паразитами сельскохозяйственных
растений, поражают древесину.
Рис. 19. Базидиомицеты
1 - холобазидии; 2, 3,4- гетеробазидии; 5 - склеробазидия.
Несовершенные грибы - очень большая группа грибов, обладающих многоклеточным мицелием, но не имеющих ни сумчатого, ни базидиального сиороношения, а лишь имеющих конидии. Размножаются исключительно бесполовым путем при помощи конидий. К этому классу относится порядок Hyphomycetales.
Среди гифомицетов имеются возбудители фузарио-токсикоза («пьяный хлеб») и интоксикаций у человека и животных.
Грибы принимают активное участие в процессах трансформации основных биогенных соединений и играют огромную роль в процессах почвообразования. Они начинают процессы разрушения наиболее стойких органических соединений углерода, таких, как лигнин, хитин, гумус, клетчатка. В процессе жизнедеятельности грибы выделяют ферменты, органические кислоты, витамины, антибиотики и другие биологически активные вещества, влияющие на развитие микроорганизмов и высших растений.
Грибы обладают большой устойчивостью к неблагоприятным факторам окружающей среды и могут активно размножаться в кислой и щелочной среде, в условиях высокой концентрации солей и недостатка влаги.