

3.Разновидности биохимического этапа.
C4- путь фотосинтеза.


Существует большая группа растений (500 видов покрытосеменных), у которых первичными продуктами фиксации СО2 и восстановления являются четырехуглеродные соединения. Их называют С4-растениями. Основы для биохимического исследования фотосинтеза у _С4-растений были заложены в работах Л. А. Незговоровой (1956-1957), К. С. Карпилова и И. А. Тарчевского (1960-1963). В 1966 г. Хетч и Слэк (австрал.) предложили законченную схему цикла темповых реакций у С4-растений, которая получила название цикла Хетча и Слэка.
К С4-растениям относится ряд культурных растений преимуественно тропического и субтропического происхождения: кукуруза, просо, сорго, сахарный тростник и многие злостные сорняки - свинорой, сыть округлая, ежовник крестьянский, просо куриное, просо крупное, гумай (сорго алепское), щирица, щетинник и др. Как правило, это высокопродуктивные растения, устойчиво осуществляющие фотосинтез при значительных повышениях температуры и в засушливых условиях.
Для листьев С4-растений характерно анатомическое строение кранц-типа (от нем. Kranz - венок, корона), т. е. наличие явно отличающихся друг от друга фотосинтезирующих клеток двух типов, которые располагаются концентрическими кругами: радиально расположенные вокруг проводящих пучков клетки обкладки и основной мезофилл (рис. 40).
Клетки обкладки проводящего пучка содержат крупные, лишенные гран (агранальные) хлоропласты. В клетках мезофилла находятся более мелкие гранальные хлоропласты. Эти два типа клеток физиологически не равноценны и специализируются на выполнении разных звеньев фотосинтеза.
C4-цикл можно разделить на две стадии: карбоксилирование (в клетках мезофилла) и декарбоксилирование и синтез углеводов (в клетках обкладки проводящих пучков). Общим для всех С4-растений является то, что карбоксилированию подвергается фосфоенолпировиноградная кислота (ФЕП) при участии ФЕП-карбоксилазы и образуется щавелевоуксусная кислота (ЩУК), которая восстанавливается до яблочной кислоты или аминируется с образованием аспарагиновой кислоты.
ЩУК, яблочкая и аспарагиновая кислоты являются C4 соединениями.
По способу декарбоксилирования при участии НАДФ-Н или НАД-малатдегидрогеназы (МДГ), называемой также малик-энзимом и яблочным ферментом) или ФЭП-карбоксикиназы (ФЕП-КК) у С4-растений можно выделить три группы: НАДФ-МДГ, НАД-МДГ и ФЕП-КК-типы соответственно.
У НАДФ-МДГ-растений главными метаболитами, вовлеченными в обмен между клетками, являются малат и пируват (ПВК), у НАД-МДГ-растений - аспартат и аланин и у ФЕП-КК-растений - аспартат и ФЕП.
Важнейшие с-х культуры (кукуруза, сорго, сахарный тростник) и такие сорняки, как сыть, ежовник, щетинник, гумай относятся к НАДФ-МДГ-типу.
Рассмотрим С4-цикл восстановления СО2 на примере этих растений (рис.).
СО2, поступающий в лист через устьица, попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с ФЕП, образуя ЩУК(оксалоацетат).
Затем в хлоропластах ЩУК восстанавливается до яблочной кислоты (малата) за счет НАДФН, образующегося в ходе световой фазы фотосинтеза; ЩУК в присутствии NH4 может превращаться также в аспартат.
Опыты с радиоактивной меткой (14С) показали, что после освещения растений в течение 1 с более 90 % радиоактивности обнаруживается в составе С4-кислот.
Затем малат переносится в хлоропласты клеток обкладки сосудистого пучка, где он подвергается окислительному декарбоксилированию, продуктом которого является ПВК. Последняя снова диффундирует в мезофилл, где при участии АТФ, образованной в световой фазе, происходит регенерация ФЕП, после чего цикл карбоксилирования повторяется с участием новой молекулы СО2. Образовавшиеся в результате окислительного декарбоксилирования малата СО2 и НАДФ-Н поступают в цикл Кальвина, что приводит к образованию ФГК и других продуктов, свойственных С3-растениям. Следовательно, именно клетки обкладки выполняют роль основной ассимилирующей ткани, поставляющей сахара в проводящую систему. Клетки мезофилла выполняют вспомогательную функцию - подкачку СО2 для цикла Кальвина.
Таким образом, С4-путь обеспечивает более полное усвоение СО2, что особенно важно для тропических растений, где основным лимитирующим фактором фотосинтеза является концентрация СО2. Эффективность усвоения СО2 С4-растениями увеличивается также за счет подачи НАДФ-Н в хлоропласты клеток обкладки. Эти хлоропласты имеют агранальное строение и специализируются на темповой фазе фотосинтеза, здесь практически не происходит НЦФФ. На один агранальный хлоропласт в среднем приходится 8-10 гранальных хлоропластов, осуществляющих первичную фиксацию СО2 и НЦФФ. Такая компартментация процессов и кооперация функционирования тканей обеспечивают повышение продуктивности растений и позволяют накапливать СО2 в органических кислотах для осуществления фотосинтеза даже при закрытых устьицах в наиболее жаркое время дня. Это сокращает потери воды на транспирацию. Эффективность использования воды С4-растениями вдвое выше, чем у С3-растений.
В 1965 г. было обнаружено, что первыми продуктами фотосинтеза у сахарного тростника (растение тропиков) являются органические кислоты, в состав которых входят 4 атома углерода (яблочная, щевелевоуксусная и аспарагиновая), а не 3С-кислота (фосфоглицериновая), как у большинства растений умеренного климата. С тех пор выявили множество растений, в основном тропических и субтропических, и имеющих порой важное экономическое значение, у которых фотосинтез протекает по такому же пути. Они получили название С4-растений. Примерами могут служить кукуруза, сорго, сахарный тростник и просо. Растения, в которых первым продуктом фотосинтеза является трехуглеродная фосфоглицериновая кислота, носят название С3-растения.
Биохимию именно С3-растений мы рассматривали до сих пор в этой главе.
В 1966 г. австралийские исследователи Хэтч и Слэк (Hatch, Slack) показали, что С4-растения значительно эффективнее, чем Сграстения, поглощают диоксид углерода: они способны удалять СО: из экспериментальной атмосферы вплоть до концентрации 0,1 части на миллион, тогда как для С3-растений это значение составляет 50—100 Ч*млн-1. Хэтч и Слэк описали новый путь метаболизма углерода у Сграсгений, получивший название путь Хэтча—Слэка. Далее мы изучим этот процесс на примере типичного Сграстения — кукурузы.
У С4-растений листья имеют характерную особенность строения: вокруг каждого проводящего пучка у них расположены два ряда клеток. У клеток внутреннего кольца — обкладки проводящего пучка — хлоропласты по своей форме отличаются от хлоропластов клеток мезофилла внешнего кольца. Различие в строении хлоро-пластов у С4-растений получило название диморфизма (наличие двух форм). На рисeyrt, Ли Б показана так называемая «кранц-анатомия» (Kranz — корона, ореол; таким образом описываются те два ряда клеток, которые окружают проводящие пучки и имеют в сечении вид двух колец). Биохимические превращения, протекающие в этих клетках, описаны ниже.
Путь Хэтча—Слэка предназначен для транспортировки диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящих пучков. Из клеток обкладки проводящих пучков диоксид углерода высвобождается и поступает в обычный С3-путь фотосинтетических превращений.
Захват (фиксация) диоксида углерода в клетках мезофилла.
Диоксид углерода фиксируется в цитоплазме клеток мезофилла. Механизм этого процесса приведен в следующем уравнении: Акцептором диоксида углерода служит фосфоенолпируват (ФЕП) вместо РиБФ у С3-растений, а вместо фермента РиБФ-карбоксилазы у С4-растений участвует фермент ФЕП-карбоксилаза. Фермент ФЕП-карбоксилаза работает значительно более эффективно, чем фермент С3-растений по двум причинам. Во-первых, ФЕП-карбоксилаза обладает большим сродством к диоксиду углерода, а во-вторых, ее работа не подвергается конкурентному ингибированию кислородом. Образовавшаяся щавелево-уксусная кислота далее превращается в малат, 4С-кислоту.
Малатный обходной путь (шунт)
Пройдя через плазмодесмы в клеточных стенках, малат попадает в хлоропласты клеток обкладки проводящего пучка, где он, соединяясь с диоксидом углерода, превращается в пируват (ЗС-кислоту). При этом выделяется водород, который используется для восстановления НАДФ. Обратите внимание, что в клетки мезофилла диоксид углерода и водород поступают извне, а затем в клетках обкладки проводящего пучка они вновь удаляются. Суммарным эффектом этих процессов является перемещение диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящего пучка.
Регенерация акцептора диоксида углерода
Пируват возвращается в клетки мезофилла, где в результате присоединения фосфатной группы от АТФ используется для регенерации ФЕП. На это расходуется энергия двух высокоэнергетических фосфатных связей.
CAM- путь фотосинтеза
С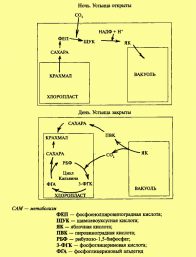 уществует
еще одна группа растений, имеющая
особенности в осуществлении первичной
фиксации С02. Это суккуленты, в частности
кактусы и растения семейства Толстянковые
(Crassulaceae). Поэтому этот тип фотосинтеза
получил название Crassulaceae acid metabolism или
САМ-метаболизм (САМ-путь). При
функционировании САМ-пути устьица
растений закрыты днем, когда интенсивность
транспирации максимальна, и открыты
ночью, когда потери воды минимальны.
Это предохраняет растения от излишней
потери воды, повышает их устойчивость
к засухе. Химизм фиксации С02 при САМ-пути
сходен с С4-путем, однако если у С4-растений
фотосинтез разделен в пространстве, то
при САМ-пути — во времени. У САМ-растений
фиксация С02 происходит в ночное время,
когда устьица открыты. С02 поступает в
клетки листа и в цитоплазме при участии
ФЕП-карбоксилазы присоединяется к
фосфоенолпировиноградной кислоте (ФЕП)
с образованием органических кислот
(щавелевоуксусной кислоты). ЩУК
восстанавливается до яблочной кислоты,
которая накапливается в вакуолях клеток
листа. Реакция идет при участии фермента
малатдегидрогеназы (малик-фермент).
Днем на свету, когда устьица закрыты,
яблочная кислота транспортируется из
вакуолей в цитоплазму, где происходит
декарбоксилирование ЩУК с образованием
С02 и пировиноградной кислоты. С02
передвигается в хлоропласта и с помощью
РБФ-карбоксилазы/оксигеназы вступает
в цикл Кальвина. Таким образом, в клетках
этих растений содержится как
ФЕП-карбоксилаза, так и
РБФ-карбоксилаза/оксигеназа. Синтезируемые
в дневные часы в цикле Кальвина углеводы
используются как источники метаболитов
(например, ФЕП), необходимых для повторения
цикла.
уществует
еще одна группа растений, имеющая
особенности в осуществлении первичной
фиксации С02. Это суккуленты, в частности
кактусы и растения семейства Толстянковые
(Crassulaceae). Поэтому этот тип фотосинтеза
получил название Crassulaceae acid metabolism или
САМ-метаболизм (САМ-путь). При
функционировании САМ-пути устьица
растений закрыты днем, когда интенсивность
транспирации максимальна, и открыты
ночью, когда потери воды минимальны.
Это предохраняет растения от излишней
потери воды, повышает их устойчивость
к засухе. Химизм фиксации С02 при САМ-пути
сходен с С4-путем, однако если у С4-растений
фотосинтез разделен в пространстве, то
при САМ-пути — во времени. У САМ-растений
фиксация С02 происходит в ночное время,
когда устьица открыты. С02 поступает в
клетки листа и в цитоплазме при участии
ФЕП-карбоксилазы присоединяется к
фосфоенолпировиноградной кислоте (ФЕП)
с образованием органических кислот
(щавелевоуксусной кислоты). ЩУК
восстанавливается до яблочной кислоты,
которая накапливается в вакуолях клеток
листа. Реакция идет при участии фермента
малатдегидрогеназы (малик-фермент).
Днем на свету, когда устьица закрыты,
яблочная кислота транспортируется из
вакуолей в цитоплазму, где происходит
декарбоксилирование ЩУК с образованием
С02 и пировиноградной кислоты. С02
передвигается в хлоропласта и с помощью
РБФ-карбоксилазы/оксигеназы вступает
в цикл Кальвина. Таким образом, в клетках
этих растений содержится как
ФЕП-карбоксилаза, так и
РБФ-карбоксилаза/оксигеназа. Синтезируемые
в дневные часы в цикле Кальвина углеводы
используются как источники метаболитов
(например, ФЕП), необходимых для повторения
цикла.
Осуществление фотосинтеза по такому пути позволяет растениям максимально экономить воду и поддерживать процесс фотосинтеза в условиях острого водного дефицита. Однако САМ-путь не может обеспечить высокой продуктивности растений, поэтому данные растения медленно растут и не могут конкурировать с С3- и С4-растениями при менее экстремальных условиях. Возможна смена путей фиксации С02. При достаточном количестве воды растения с САМ-метаболизмом могут переходить на С3-путь. Так, показана способность Mesembryanthemum crystallinum L. (хрустальная травка) менять направленность метаболических процессов, переходя с С3-пути фотосинтеза на САМ в условиях действия различных стрессоров: водного, солевого, температурного. В научной литературе имеются указания о наличии и других путей связывания С02. Однако они еще недостаточно изучены. Важно также подчеркнуть, что на всех этапах фотосинтетического цикла промежуточные продукты могут претерпевать различные превращения в зависимости от условий среды. Именно это является основой для образования разнообразных продуктов фотосинтеза.
С2- путь фотосинтеза.
Фотодыхание (гликолатный путь, С2-фотосинтез) — стимулируемое светом выделение углекислого газа и поглощение кислорода у растений преимущественно с С3-типом фотосинтеза. Также под фотодыханием понимают биохимический путь, связанный с регенерацией одной молекулы 3-фосфоглицериновой кислоты (С3) из двух молекул гликолевой кислоты (С2) и лежащий в основе вышеописанного газообмена. Наличие биохимического механизма фотодыхания обусловлено значительной оксигеназной активностью РуБисКО, ключевого фермента цикла Кальвина.
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) - CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
П уть
восстановления СО2 при фотосинтезе М.
Кальвин изучал при относительно высоких
концентрациях СО2 .
При более низких концентрациях СО2 ,
приближающихся к атмосферным (0,045 %), в
хлоропласте образуется значительное
количество фосфогликолевой кислоты: в
условиях низкого парциального давления
СО2 и высокого парциального давления
О2 РДФ-карбоксилаза может присоединять
к рибулозо-1,5-дифосфату кислород – может
выполнять не только карбоксилирующую,
но и оксигенерирующую функции (оксигеназа).
Присоединение кислорода к молекуле
рибулозо-1,5-дифосфата приводит к тому,
что вместо 2 молекул ФГК образуется 1
молекула ФГК и 1 молекула фосфогликолата.
1 молекула фосфогликолата во время
транспорта через мембрану хлоропласта
дефосфорилируется при участии
фосфогликолатфосфатазы и превращается
в гликолат, который поступает в специальную
органеллу-пероксисому
Цикл превращения гликолата (первичного
продукта) в триозофосфат (ФГК)- гликолатный.
уть
восстановления СО2 при фотосинтезе М.
Кальвин изучал при относительно высоких
концентрациях СО2 .
При более низких концентрациях СО2 ,
приближающихся к атмосферным (0,045 %), в
хлоропласте образуется значительное
количество фосфогликолевой кислоты: в
условиях низкого парциального давления
СО2 и высокого парциального давления
О2 РДФ-карбоксилаза может присоединять
к рибулозо-1,5-дифосфату кислород – может
выполнять не только карбоксилирующую,
но и оксигенерирующую функции (оксигеназа).
Присоединение кислорода к молекуле
рибулозо-1,5-дифосфата приводит к тому,
что вместо 2 молекул ФГК образуется 1
молекула ФГК и 1 молекула фосфогликолата.
1 молекула фосфогликолата во время
транспорта через мембрану хлоропласта
дефосфорилируется при участии
фосфогликолатфосфатазы и превращается
в гликолат, который поступает в специальную
органеллу-пероксисому
Цикл превращения гликолата (первичного
продукта) в триозофосфат (ФГК)- гликолатный.
В растительных клетках, содержащих хлоропласты, помимо С3- и С4-путей фотосинтеза, осуществляется также фотодыхание, т. с. активируемый светом процесс высвобождения СО2, и поглощения О2,, который значительно отличается от «темнового» дыхания митохондрий. Так как при этом первичным продуктом является гликолевая кислота, то этот путь получил название гликолатного. У .некоторых С3-растений с малой эффективностью фотосинтеза интенсивность фотодыхания может достигать 50% от интенсивности фотосинтеза.
Фотодыхание у C3-растений обычно усиливается при низком содержании С02 и высоких концентрациях 02. В этих условиях РДФ-карбоксилаза в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая затем дефосфорилируется в гликолевую кислоту (рис.). Молекулы С02 и 02 конкурируют между собой в каталитическом центре РДФ-карбоксилазы: при относительно высоких концентрациях С02 и низких 02 преобладает карбоксилирование, тогда как высокие концентрации 02 и низкое содержание СО2, благоприятствуют окислению, а следовательно, и образованию фосфогликолевой кислоты. В том же направлении действует и повышение температуры.
Фотодыхание осуществляется в результате взаимодействия трех органелл — хлоропластов. пероксисом и митохондрий (рис.). Гликолат из хлоропласта поступает в пероксисому и там окисляется гликолатоксидазой до глиоксилата. Возникающая перекись водорода устраняется каталазой пероксисомы. Глиоксилат аминируется, превращаясь в глицин, причем в качестве донора аминогруппы функционирует глутамат.
Глицин транспортируется в митохондрию. Здесь из двух молекул глицина образуется серии и освобождается С02. Теперь цикл замыкается: серин может снова поступать в пероксисому и там передать свою аминогруппу на пируват. При этом из пирувата возникает аланин, из серина — гидроксипируват, который сразу после этого восстанавливается вглицерат. Затем глицерат может снова попасть в хлоропласты и благодаря фосфорилированию включиться в цикл Кальвина.
Последовательность реакций фотодыхания не обязательно образует цикл. Гликолатный путь С3-растений может завершиться в митохондриях. Конечными продуктами в этом случае являются серин и С02. Освобождение С02 объясняет, почему нетто-фотосинтез (чистая продуктивность) при интенсивном фотодыхании снижается (рис.).
У С4-расгений С02, выделяющийся в результате фотодыхания, перехватывается в клетках мезофилла, где из ФЕП и С02 образуются оксалоацетат и малат. Затем малат «отдает» свой С02 хлоропластам обкладки, где функционирует цикл Кальвина. В связи с этим становится понятным высокий нетто-фотосинтез С4-растений.
С3- путь фотосинтеза.
С3-фотосинтез, цикл Кальвина.
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6H12O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий: .
Первая фаза — карбоксилирование. Эта реакция катализируется специфическим для процесса фотосинтеза ферментом рибулозобисфосфат-карбоксилазой/ оксигеназой (сокращенно РБФ-карбоксилаза/оксигеназа), в научной литературе последних лет чаще встречается под названием Rubisco, от ribulosobiphosphatecarboxylase/oxygenase. Впервые Rubisco был выделен и очищен в 1955 году. Особенностью фермента является то, что катализируемая им реакция является самой медленной стадией в цикле фиксации углекислоты. В листьях Rubisco содержится в больших количествах и является основной фракцией белка хлоропластов. Более того, высказывается мнение, что это самый распространенный белок-фермент на земном шаре. В активное состояние фермент переходит при освещении хлоропластов. Уже отмечалось, что формирование этого фермента происходит под контролем двух геномов: большие субъединицы (54 кДа) кодируются в ядре, синтезируются в цитоплазме; малые — кодируются и синтезируются в хлоропласте. При взаимодействии РБФ с С02 образуется сначала промежуточное нестойкое шестиуглеродное соединение, которое затем распадается на две молекулы ФГК. Образовавшаяся ФГК — это органическая кислота, и ее энергетический уровень ниже уровня Сахаров. Поэтому это соединение не может непосредственно превращаться в углеводы. Необходимо превращение его в трехуглеродный сахар — фосфоглицериновый альдегид (ФГА).
Вторая фаза — восстановление. Дальнейшие превращения ФГК требуют участия продуктов световой фазы фотосинтеза: АТФ и НАДФН + Н+. Реакция идет в два этапа. Прежде всего, происходит реакция фосфорилирования 3-ФГК. Донором фосфатной группы является АТФ. АТФ требуется здесь в качестве дополнительного источника энергии. Образуется 1,3-дифосфоглицериновая кислота. Реакция катализируется ферментом фосфоглицерокиназой:

фосфорилирования 3-ФГК
Образовавшееся в этой реакции соединение — дифосфоглицериновая кислота — обладает более высокой реакционной способностью, содержит макроэргическую связь, полученную от АТФ. Затем карбоксильная группировка этого соединения восстанавливается до альдегидной с помощью триозофосфатдегидрогеназы, коферментом которой служит НАДФ.
Образовавшийся ФГА является по уровню восстановленным углеводом. Это соединение вступает в две последние фазы. Пять молекул ФГА используется на регенерацию акцептора РБФ для того, чтобы фиксация С02 могла снова осуществляться. Оставшаяся шестая молекула вступает в фазу «синтеза продуктов», где превращается в более сложные соединения (углеводы, аминокислоты и др.).
Третья фаза — регенерация. В процессе регенерации акцептора используется пять молекул ФГА, в результате чего образуются три молекулы рибулезо-5-фос-фата. Этот процесс идет через образование 4-, 5-, 6-, 7-углеродных соединений. Прежде всего, первая молекула ФГА изомеризуется до фосфодиоксиацетона. Процесс катализируется ферментом триозофосфатизомеразой. От ФДФ отщепляется фосфат, и ФДФ превращается во фруктозо-6-фосфат (ф-6-Ф). Далее от Ф-6-Ф (С6) отщепляется 2-углеродный фрагмент (—СО—СН2ОН), который переносится на следующую (третью) триозу. Это транскетолазная реакция идет при участии фермента транскетолазы. В результате образуется первая пентоза (С5)-рибулезофосфат. От Ф-6-Ф остается 4-углеродный сахар эритрозофосфат фосфорилирование. Для этого используются три молекулы АТФ, образовавшиеся в результате световых реакций. Все реакции, входящие в цикл Образовавшийся триозофосфат (ФГА) вступает в четвертую стадию темновых реакций — стадию образования продуктов фотосинтеза. (С4), который конденсируется с четвертой триозой с образованием седогептулезодифосфата (С7). После отщепления фосфата седогептулезодифосфат превращается в седогептулезофосфат. Далее снова происходит транскетолазная реакция, в результате которой от седогептулезофосфата отщепляется 2-углеродный фрагмент, который переносится на пятую триозу Образуются еще две молекулы рибулезофосфата. Таким образом, в результате рассмотренных реакций получаются 3 молекулы рибулезофосфата. При прохождении двух циклов из 12 молекул образовавшегося ФГА две молекулы выходят из них, образуя одну молекулу фруктозодифосфата (ФДФ). Из двух молекул фруктозодифосфата (Ф-1, 6-диФ) образуются фруктозо-6-фосфат (ф-6-Ф) и глюкозо-1-фосфат (Г-1-Ф). Глюкозо-1-фосфат, взаимодействуя с уридинтрифосфатом (УТФ), дает уридиндифосфоглюкозу (УДФГ). В свою очередь УДФГ, реагируя с Ф-6-Ф, дает сахарозофосфат. По-видимому, именно сахароза является первым свободным сахаром, образующимся в процессе фотосинтеза. Из сахарозы образуются нефосфорилированные моносахара (глюкоза и фруктоза). Крахмал образуется из аденозиндифосфоглюкозы (АДФГ) или уридиндифосфоглюкозы (УДФГ), процесс катализируется ферментом амилосинтетазой. Среди первых продуктов фотосинтеза обнаружены аминокислоты. По-видимому, ФГК, образовавшаяся на первом этапе цикла Кальвина, может превращаться в пировиноградную кислоту. Этот процесс идет особенно интенсивно при недостатке НАДФН, из-за чего задерживается преобразование ФГК в ФГА (обычный путь в цикле Кальвина). Пировиноградная кислота в присутствии NH3 дает аминокислоту аланин. Показано, что скорость включения 14С02 в аланин в клетках хлореллы при некоторых условиях может даже превышать скорость ее включения в сахарозу. Из пировиноградной кислоты может образоваться еще ряд органических кислот (в цикле Кребса).
Образовавшиеся органические кислоты в процессе аминирования или переаминирования дают аминокислоты. Сам по себе синтез аминокислот еще не означает образование белков. Однако было показано, что между этими двумя процессами имеется прямая связь. Так, под влиянием освещения синими лучами (458—480 нм) усиливается фотосинтетическое образование, как аминокислот, так и белков. В присутствии ингибиторов синтеза белка действие синего света не проявляется. Наконец, из промежуточных продуктов цикла Кальвина могут образовываться жиры, липиды и другие продукты. Состав продуктов, образующихся при фотосинтезе, может быть определен исходя из величин фотосинтетического коэффициента. Под фотосинтетическим коэффициентом понимается отношение выделенного в процессе фотосинтеза кислорода к поглощенному С02. Если в процессе фотосинтеза образуются углеводы, то, согласно приведенному суммарному уравнению, фотосинтетический коэффициент должен быть равен единице: 602/6С02 = 1. При образовании соединений, более восстановленных (содержащих меньше кислорода) по сравнению с углеводами, фотосинтетический коэффициент должен быть больше единицы. В случае образования белков фотосинтетический коэффициент равен 1,25, в случае жира — 1,44. Средняя величина фотосинтетического коэффициента для 27 видов растений оказалась равной 1,04. Расчеты показали, что такая величина фотосинтетического коэффициента указывает на образование наряду с углеводами некоторого количества белка (примерно 12%). Установлено, что величина фотосинтетического коэффициента меняется в зависимости от условий. Преобладание синих лучей над красными приводит к увеличению доли образующихся белков, тогда как красный свет благоприятствует образованию углеводов (Н.П. Воскресенская). Усиление снабжения растений азотом, естественно, также приводит к повышению фотосинтетического коэффициента и увеличению первичного синтеза белка. Имеются данные, что на качество продуктов фотосинтеза оказывает влияние интенсивность освещения. При высокой освещенности образуется больше углеводов, а при пониженной — аминокислот.