

Водный режим. Минеральное питание
ЗНАЧЕНИЕ ВОДЫ В ЖИЗНЕДЕЯТЕЛЬНОСТИ РАСТЕНИЙ
Вода является основной составной частью растительных организмов. Ее содержание доходит до 95% от массы организма, и она участвует прямо или косвенно во всех жизненных проявлениях.
Вода — это та среда, в которой протекают все процессы обмена веществ.
Она составляет основную часть цитоплазмы, поддерживает ее структуру, устойчивость входящих в состав цитоплазмы коллоидов, обеспечивает определенную конформацию молекул белка. Высокое содержание воды придает содержимому клетки (цитоплазме) подвижный характер.
Вода — непосредственный участник многих химических реакций. Все реакции гидролиза, многочисленные окислительно-восстановительные реакции идут с участием воды, например реакции цикла Кребса.
Вода является донором электронов и протонов при фотосинтезе.
Передвижение веществ по растению в сосудах ксилемы и ситовидных трубках осуществляется в водной среде.
Терморегуляция и защита тканей от резких колебаний температуры.
Поддержание упругого состояния клеток и защита от механических повреждений.
Согласно современным представлениям, вода в клетке представляет собой сложную гетерогенную систему, состоящую из:
1) жидкой фазы;
2) гидратно-связанной;
3) гидрофобно-стабилизированной (главным образом в мембранах);
4) пространственно стабилизированной (в капиллярных промежутках).
Что касается гидратной воды, то различают два типа гидратации:
1) притяжение диполей воды к заряженным частицам (как к ионам минеральных солей, так и к заряженным группам белка СОО- и NН4+);
2) образование водородных связей с полярными группами органических веществ —между водородом воды и атомами О или N. Вода в растении находится как в свободном, так и в связанном состоянии. Свободной называют воду, сохранившую все или почти все свойства чистой воды. Свободная вода легко передвигается, вступает в различные биохимические реакции, испаряется в процессе транспирации и замерзает при низких температурах. Связанная вода имеет измененные физические свойства главным образом вследствие взаимодействия с неводными компонентами.
Воду, гидратирующую коллоидные частицы (прежде всего белки), называют коллоидно-связанной, а растворенные вещества (минеральные соли, сахара, органические кислоты и др.) — осмотически-связанной. Некоторые исследователи считают, что вся вода в клетке в той или иной степени связана. Физиологи условно понимают под связанной водой ту, которая не замерзает при понижении температуры до —10 °С. Важно отметить, что всякое связывание молекул воды (добавление растворенных веществ, гидрофобные взаимодействияи др.) уменьшает их энергию. Именно это лежит в основе снижения
водного потенциала клетки по сравнению с чистой водой.
ЗАКОНОМЕРНОСТИ ПОГЛОЩЕНИЯ ВОДЫ
В настоящее время говорят о двух механизмах поступления: осмотическом и коллоидно-химическом. Осмотический механизм – в основе лежит осмос – причиной которого является разность концентраций растворов по обе стороны полупроницаемой мембраной. Более концентрированный раствор – клеточный сок, менее концентрированный – в свободном пространстве клеточной стенке. Однако осмос нельзя считать просто односторонней диффузией, не зависящей от обмена веществ, для него нужна энергия. Она тратится на создание разных концентраций по обе стороны мембраны – на активный транспорт растворенных веществ в вакуоль, на образование веществ (продуктов метаболизма), сохранение избирательной проницаемости. Осмотическое поступление воды приводит к возникновению гидростатического (тургорного) давления. Разница между осмотичечским давлением клеточного сока и противодавлением клеточной стенки определяет поступление воды в клетку в каждый данный момент.
Коллоидно-химический механизм. Осмотическое поступление воды связано в основном с растворенными веществами клеточного сока, т.е. с вакуолью. Однако около 80% сухой массы протопласта составляют высокомолекулярные соединения: белки, полисахариды, нуклеиновые кислоты, нерастворимые в воде. Они тоже влияют на поступление воды в клетку. Имея отрицательно и положительно заряженные группы атомов, гидрофильные коллоиды притягивают диполи воды. Молекулы воды могут удерживаться около молекулы полимера и с помощью водородных связей, между атомами водорода и кислорода. Поглощение жидкости высокомолекулярным веществом, сопровождаемое увеличением его объема, называется набуханием. Набухание может рассматриваться как особый вид диффузии, так как движение воды идет по градиенту концентрации. Силу набухания называют матричным потенциалом. Набухание участвует в поступлении воды в цитоплазму и в клеточную стенку. Сухие семена поглощают воду исключительно с помощью набухания, особенно сильно бобовые с большим содержанием белков.
Особенностью растений является наличие аквапоринов – специальных белков, которые встраиваясь в мембраны, образуют в билипидном слое водные каналы или поры, ускоряя диффузию воды. Внутренняя поверхность образованной поры гидрофильна, и вода легко проходит через мембрану. Образование и активность функционирования аквапоринов контролируется различными регуляторными системами и зависят от внешних условий.
ПЕРЕДВИЖЕНИЕ ВОДЫ ПО РАСТЕНИЮ
Вода, поступившая в клетки корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно независимых друг от друга объемов, по которым передвигается вода,— апопласта и симпласта. Апопласт — это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт — это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая — по другую сторону клеток эндодермы и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам. Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту. Однако, для того чтобы попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Вода устремляется через эту мембрану в сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы. Как уже упоминалось, по вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса, это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация и водный потенциал становится более отрицательным. Предполагается, что в результате активного (с затратой энергии) поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикла), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды. Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид. Вне ксилемы движется всего от 1 до 10% общего потока воды. Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры — отличительной чертой растении, устойчивых к засухе.
Вода передвигается от клетки к клетке благодаря градиенту водного потенциала. По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам, где сопротивление значительно меньше.
По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Вызванное транспирацией движение воды по растению называют транспирационным током.
Непрерывно двигаясь по растению, вода доставляет клеткам вещества, необходимые для их функционирования, и одновременно уносит из клеток часть веществ, в том числе и ненужных. Благодаря транспорту воды осуществляется связь между клетками, органами, обеспечивающая нормальное протекание всех физиологических процессов.
Передвижение минеральных веществ
Ближний транспорт в корне. Поглощенные клетками эпиблемы корня ионы и продукты их ассимиляции могут двигаться к сосудам центрального цилиндра двумя путями: по симпласту и апопласту. Корневой волосок это главный вход в симпласт, из него или других клеток эпиблемы поглощенный ион переходит из одной клетки коры в другие по плазмодесмам. Внутри симпласта ионы и их органические производные передвигаются от клетки к клетке по плазмодесмам без преодоления мембран со скоростью 1,5-2,0 см/ч. Войдя в протопласт, ион может затем попасть внутрь эндоплазматического ретикулума. Так как в состав каждой плазмодесмы входит его канал, то попавший в него ион может транспортироваться по симпласту из клетки в клетку, не выходя в цитозоль.
Механизм транспорта ионов по симпласту до конца не выяснен. Предполагают, что это диффузия и массовый ток веществ из клетки в клетку по градиенту концентрации. Этот концентрационный градиент создают корневые волоски, активно поглощая вещества. Допускается возможность и поверхностной миграции ионов вдоль плазмодесм.
Вакуоли клеток поддерживают концентрацию данного иона в симпласте постоянной. Если во внешней среде содержится оптимальное количество иона, то вакуоль не оказывает влияния на его транспорт по симпласту, если недостаток или нет совсем этого иона – то он поступает из вакуоли в цитозоль и потом в симпластический путь, если избыток – происходит его накопление в вакуоли. Поглощение ионов вакуолями снижает их концентрацию в симпласте и обеспечивает создание градиента концентрации, необходимого для транспорта ионов по симпласту. Корень способен выполнять распределительную функцию, направляя в побеги дефицитные элементы питания и задерживая в вакуолях избыточные или вредные.
Основным путем апопластного транспорта являются рыхлые первичные клеточные стенки. Однако в апопласте свобода передвижения ограничена и объясняется тем, что карбоксильные клеточной стенки несут отрицательный заряд, поэтому катионы легко адсорбируются на поверхности и могут мигрировать далее по ней в глубь корня, анионы же отталкиваются одноименными зарядами клеточной стенки и поэтому проникают в апопласт труднее. В результате неодинакового проникновения катионов и анионов через клеточные стенки внутри корня создается избыток катионов, а следовательно положительный потенциал, препятствующий их дальнейшему проникновению.
Между апопластом и симпластом может идти обмен ионами. Он происходит в области пор, где вторичные стенки отсутствуют. Передвигаясь по апопласту, вещества, как и вода, встречают меньшее сопротивление, чем при движении через плазмалемму.
Ионы, как и молекулы воды, по апопласту могут двигаться до эндодермы. В эндодерме происходит обязательный переход ионов с апопластного на симпластный путь. Это переключение позволяет регулировать ток ионов. Основным механизмом движения по апопласту также считают диффузию и массовый ток. Особенностью апопластного пути – зависимость движения ионов от воды поэтому его называют водозависимым.
В целом главным для транспорта многих ионов является симпластный путь, при этом некоторые из них особенно азот и фосфор во время транспорта включаются в химические соединения.
Ионы поступают в клетки ризодермы из почвенного раствора и благодаря контактному обмену Н+, НСО-3 и анионов органических кислот на ионы минеральных веществ почвенных частиц. Контактный обмен ионов клеточной стенки ризодермы с частицами почвы осуществляется без перехода ионов в почвенный раствор. Тесный контакт обеспечивается благодаря выделению слизи корневыми волосками и отсутствию у ризодермы кутикулы и других защитных покровов. Так как адсорбированные ионы находятся в постоянном колебательном движении и занимают определенный объем - сферу колебаний, при тесном контакте поверхностей сферы колебаний двух ближайших адсорбированных ионов могут перекрываться, в результате чего осуществляется ионный обмен.
Выделяя различные вещества (углекислый газ, аминокислоты, сахара и другие), корень растения изменяет состояние питательных веществ в прикорневой зоне непосредственно, например, путем выделения СО2 (СО2 + Н2О Н+ + НСО-3: повышение растворимости фосфатов и карбонатов) и косвенно, создавая благоприятные условия для ризосферы, которая играет большую роль в превращении почвенных минералов.
Растения способны поглощать из окружающей среды практически все элементы. Однако для нормальной жизнедеятельности растительному организму необходимо лишь 19 питательных элементов. Среди них углерод (45 % сухой массы тканей), кислород (42%), водород (6,5%) и азот (1,5%) называют органогенами. Оставшиеся 5 % приходятся на зольные элементы, которые остаются в золе после сжигания растения. Содержание минеральных элементов обычно выражают в процентах от массы сухого вещества. Все элементы в зависимости от их количественного содержания в растении принято делить на макроэлементы (содержание более 0,01 %) - к ним относятся азот, фосфор, сера, калий, кальций, магний и микроэлементы (содержание менее 0,01 %): железо, марганец, медь, цинк, бор, молибден, кобальт, хлор. Ю. Либихом было установлено, что все перечисленные элементы равнозначны и полное исключение любого из них приводит растение к глубокому страданию и гибели, ни один из перечисленных элементов не может быть заменен другим, даже близким по химическим свойствам. Макроэлементы при концентрации 200-300 мг/л в питательном растворе еще не оказывают вредного действия на растение. Большинство микроэлементов при концентрации 0,1-0,5 мг/л угнетают рост растений.
Для нормальной жизнедеятельности растений должно быть определенное соотношение различных ионов в окружающей среде. Чистые растворы одного какого-либо катиона оказываются ядовитыми. Так, при помещении проростков пшеницы на чистые растворы KCL или CaCL2 на корнях сначала появлялись вздутия, а затем корни отмирали. Смешанные растворы этих солей не обладали ядовитым действием. Смягчающее влияние одного катиона на действие другого катиона называют антагонизмом ионов. Антагонизм ионов проявляется как между разными ионами одной валентности, например, между ионами натрия и калия, так и между ионами разной валентности, например, калия и кальция. Одной из причин антагонизма ионов является их влияние на гидратацию белков цитоплазмы. Двухвалентные катионы (кальций, магний) дегидратируют коллоиды сильнее, чем одновалентные (натрий, калий). Следующей причиной антагонизма ионов является их конкуренция за активные центры ферментов. Так, активность некоторых ферментов дыхания ингибируется ионами натрия, но их действие снимается добавлением ионов калия. Кроме того, ионы могут конкурировать за связывание с переносчиками в процессе поглощения. Действие одного иона может и усиливать влияние другого иона. Это явление называется синергизмом. Так, под влиянием фосфора повышается положительное действие молибдена.
ПРИМЕНЕНИЕ УДОБРЕНИЙ
В естественных биоценозах поглощенные из почвы соединения частично возвращаются с опавшими листьями, ветками, хвоей. С убранным урожаем сельскохозяйственных растений поглощенные вещества из почвы устраняются. Величина выноса минеральных элементов зависит от вида растения, урожайности и почвенно-климатических условий. Овощные культуры, картофель, многолетние травы выносят больше элементов питания, чем зерновые.
Для предотвращения истощения почвы и получения высоких урожаев сельскохозяйственных культур необходимо внесение удобрений. Сопоставляя количество элементов в почве и растении с величиной урожая Ю. Либих сформулировал закон минимума или закон ограничивающих факторов. Согласно этому закону величина урожая зависит от количества того элемента, который находится в почве в относительном минимуме. Увеличение содержания этого элемента в почве за счет внесения удобрений будет приводить к возрастанию урожая до тех пор, пока в минимуме не окажется другой элемент. Позже было установлено наличие у растений критических периодов по отношению к тому или иному минеральному элементу, то есть периодах более высокой чувствительности растений к недостатку этого элемента на определенных этапах онтогенеза. Это позволяет регулировать соотношение питательных веществ в зависимости от фазы развития и условий среды. Так, известно, что в осенний период для озимых культур не рекомендуется вносить азотные удобрения, так как они усиливают ростовые процессы, снижая устойчивость растений. В осенний период надо проводить подкормки фосфором и калием, а весной азотом.
С помощью удобрений можно регулировать не только величину урожая, но и его качество. Так, для получения зерна пшеницы с высоким содержанием белка надо вносить азотные удобрения, а для получения продуктов с высоким содержанием крахмала (например, зерна пивоваренного ячменя или клубни картофеля) необходимы фосфор и калий. Внекорневая подкормка фосфором незадолго до уборки усиливает отток ассимилятов из листьев сахарной свеклы к корнеплодам и тем самым увеличивает ее сахаристость.
Система удобрений - это программа применения удобрений в севообороте с учетом растений-предшественников, плодородия почвы, климатических условий, биологических особенностей растений, состава и свойств удобрений. Система удобрений создается с учетом круговорота веществ и их баланса в земледелии. Баланс питательных веществ учитывает поступление их в почву с удобрениями, суммарный расход на формирование урожаев и непродуктивные потери в почве. Необходимое условие функционирования системы удобрений - это предотвращение загрязнения окружающей среды вносимыми в почву химическими соединениями.
Удобрения подразделяют на минеральные и органические, промышленные (азотные, калийные, фосфорные, микроудобрения, бактериальные) и местные (навоз, торф, зола), простые (содержат один элемент питания - азотные, калийные, борные) и комплексные (содержат два или более питательных элементов). Среди комплексных удобрений выделяют сложные и комбинированные. Сложные удобрения в составе одного химического соединения содержат два или три питательных элемента, например, калийная селитра (KNO3), аммофос (NH4H2PO4) и другие. Одна гранула комбинированных удобрений включает два или три основных элемента питания в виде различных химических соединений, например, нитроаммофоска.
До посева вносят 2/3 общей нормы удобрений. Они должны обеспечить растение на весь период развития элементами питания и повысить плодородие почвы. Припосевное удобрение в виде хорошо растворимых соединений вносят малыми дозами одновременно с посевом или посадкой растений для обеспечения минерального питания молодых растений. Послепосевные внекорневые подкормки, основанные на способности листьев поглощать минеральные соли в растворе, проводятся для усиления питания растений в наиболее важные периоды их развития.
Транспирация, ее значение для растений и методы изучения.
Транспирация — процесс движения воды через растение и её испарение через наружные органы растения, такие как листья, стебли и цветки.
Значения: Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете; Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое; С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом чем интенсивнее транспирация, тем быстрее идет этот процесс.Основным транспирирующим органом является лист. Средняя толщина листа составляет 100—200 мкм. Паренхимные клетки листа расположены рыхло, между ними имеется система межклетников. Эпидермис — покровная ткань листа, состоит из компактно расположенных клеток, наружные стенки которых утолщены. Кроме того, листья большинства растений покрыты кутикулой. Удаление кутикулы во много раз повышает интенсивность испарения. Для соприкосновения листа с атмосферой имеются поры — устьица.
У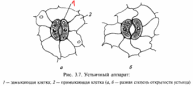 стьице —
это отверстие (щель), ограниченная двумя
замыкающими клетками. Каждая замыкающая
клетка устьица в отличие от клеток
эпидермиса имеет хлоропласта. В них
происходит фотосинтез, хотя с меньшей
интенсивностью, чем в клетках мезофилла.
Устьица — одно из оригинальных
приспособлений, обладающих способностью
открываться и закрываться в зависимости
от насыщенности замыкающих клеток
водой. Обычно устьичные отверстия
ограничены двумя замыкающими клетками,
стенки которых неравномерно утолщены.
У двудольных растений замыкающие клетки
бобовидной, или полулунной, формы, при
этом их внутренние прилегающие друг к
другу клеточные стенки более толстые,
а внешние — более тонкие.
стьице —
это отверстие (щель), ограниченная двумя
замыкающими клетками. Каждая замыкающая
клетка устьица в отличие от клеток
эпидермиса имеет хлоропласта. В них
происходит фотосинтез, хотя с меньшей
интенсивностью, чем в клетках мезофилла.
Устьица — одно из оригинальных
приспособлений, обладающих способностью
открываться и закрываться в зависимости
от насыщенности замыкающих клеток
водой. Обычно устьичные отверстия
ограничены двумя замыкающими клетками,
стенки которых неравномерно утолщены.
У двудольных растений замыкающие клетки
бобовидной, или полулунной, формы, при
этом их внутренние прилегающие друг к
другу клеточные стенки более толстые,
а внешние — более тонкие.
Кутикулярная транспирация Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации.Интенсивность кутикулярной транспирации варьирует у разных видов растений. У молодых листьев с тонкой кутикулой она может составлять около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация равна 1/10 общей транспирации. В стареющих листьях из-за повреждения кутикулы она может возрастать. Таким образом, кутикулярная транспирация регулируется главным образом толщиной и целостностью кутикулы и других защитных покровных слоев на поверхности листьев. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом.
Устьичная транспирация Основная часть воды испаряется через устьица. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно
Количественные показатели транспирации: Интенсивность транспирации – это количество, г, воды, испаряемой растением в единицу времени (ч) с единицы поверхности (дм 2 ). При определении продукционных характеристик рассчитывают ко- личество воды, израсходованной растением за весь вегетационный период, и относят его к сухой массе всего растения. Транспирационный коэффициент – это количество воды (г), расходуе- мой растением на образование 1 г сухого вещества.
Сутчный ход транспирации. У всех растений наблюдается периодичность суточного хода транспирации. У деревьев, теневых растений злаков (гидростабильные виды) испарение воды достигает максимума до наступления максимума дневной температуры. В полуденные часы транспирация падает. Вечером, при снижении дневных температур транспирация снова увеличивается. Такой ход транспирации приводит к незначительным изменениям осмотического давления и содержания воды в клетках в течение дня.
У видов, способных переносить резкие изменения содержания воды в клетках в течение дня, транспирация повышается в полдень и падает ночью (гидролабильные виды) Закрывание устьиц в полдень может быть вызвано увеличением уровня углекислого газа в листьях при повышении температуры воздуха (усиление дыхания и фотодыхания), а также возможным водным дефицитом, возникающим в тканях при высоких температурах и низкой влажности воздуха. Это приводит к повышению концентрации АБК и закрыванию устьиц.
На поступление воды в растение оказывают влияние внешние условия.
1. Температура. Поступление воды в растение зависит от температуры. С понижением температуры скорость поступления воды сокращается. Это может происходить в результате следующих причин:
а) повышается вязкость воды и снижается ее подвижность;
б) Тормозится рост корней;
в) Уменьшается скорость метаболических процессов;
2. Снижение аэрации почвы (повышение углекислого газа) Повышение концентрации углекислого газа приводит к повышению вязкости воды и снижает проницаемость цитоплазмы.
3. Содержание воды в почве, концентрация почвенного раствора. Вода поступает в корень, если водный потенциал корня ниже, чем водный потенциал почвы. На засоленных почвах или на почвах, где концентрация почвенного раствора очень высокая, водный потенциал почвы ниже. Поэтому вода начнет выделяться из корня. У растений, произрастающих на этих почвах - галофитах, в процессе эволюции выработался такой приспособительный признак как высокая
концентрация клеточного сока. Это обуславливает более низкий водный потенциал клеточного сока, вследствие чего вода из почвенного раствора поступает в корни.
Методы транспирации
Чаще всего применяют или метод быстрого взвешивания, или метод инфильтрации (открытость устьиц). Первый метод связан с сильным нарушением, но все же он дает некоторые сравнительные результаты. Расчет транспирации обычно производится в миллиграммах испаряемой воды (потери массы срезанного побега) за какую-то единицу времени (час, минуту) на единицу поверхности или на сырую/сухую массу.
Расчет на сырую массу сильно меняется в течение дня. И, кроме того, надо учитывать и анатомические особенности, прежде всего выраженность в структуре листа механических тканей. Расчет транспирации на сухую массу дает так называемый транспирационный коэффициент, который указывает, сколько растение затратило воды (в граммах) на производство единицы массы сухого органического вещества (фитомассы).
Общей закономерностью является то, что при ухудшении условий транспирационный коэффициент увеличивается, т. е. растение должно «прогнать» через себя больше воды на производство фитомассы. Если мы хотим сравнить транспирацию нескольких видов сообщества, то удобно один из них выбрать за единицу (эталон) и тогда можно получить "сравнительную транспирацию". Новый метод измерения транспирации в древесных стволах в почвенно-климатических условиях хорезмской области основан на датчики теплового рассеивания. Датчики непосредственно измеряют скорость ксилемного сока, которая может быть соотнесена к транспирации дерева рассчитанной на единицу площади активного ксилема. Принцип данного метода был развит доктором Андре Граниером во Франции и был широко принят сообществом исследователей, начиная с 1996.
Величину транспирации растений также можно определить весовым или объемным методом. Массу воды, которую растение испаряет в единицу времени, определяют путем систематического взвешивания вегетационного сосуда вместе с выращиваемыми растениями. При объемном методе пользуются прибором – потометром. Для измерения фактической транспирации в естественных условиях используют почвенные испарители, или лизиметры.
Определение интенсивности транспирации при помощи торсионных весов по методу А. Л. Иванова. При определении интенсивности транспирации отрезанных листьев производят учет изменения их массы за короткие промежутки времени, обычно за 1-2 мин. Это дает возможность наблюдать транспирацию при том состоянии насыщенности водой листа, в каком он находился на растении. В основе метода лежит учет количества воды, потерянной листом за короткий промежуток времени (1-15 мин в зависимости от условий опыта) в результате транспирации. При этом изменение массы листа следует учитывать до начала его завядания.
Определение интенсивности транспирации объёмным методом (в модификации В. П. Моисеева). Основным достоинством данного метода является его точность, наглядность, возможность изучения динамики транспирации в различных условиях среды. Для выполнения работы не требуется сложной измерительной техники. Сущность метода состоит в том, что лист при помощи эластичной трубки соединяется с микропипеткой (цена деления 0,001- 0,002 мл), предварительно заполненной водой. Объем поглощенной листом воды из пипетки будет соответствовать величине транспирации.
ОСОБЕННОСТИ ПИТАНИЯ РАСТЕНИЙ АЗОТОМ
Без N не могут синтезироваться белковые в-ва, а без них не может образоваться протопласт живой клетки. В растениях 1-3% N, без достаточного кол-ва его в почве развитие растений невозможно. Низкая урожайность многих с/х культур чаще всего определяется недостатком N. В почв N содержится в виде органических соединений (белковые в-ва и аминокислоты) и минеральных соединений (аммонийные соли и соли азотной к-ты). Весь N растениями усваивается из почвы. Различают несколько видов соединений N: органические соединения (орг.N), соли аммиака (аммиачный N), соли азотной и азотистой к-т (нитратный N). Общее кол-во N в почве незначительно, в основном это органический N. Органические соединения, которые усваиваются растениями: аминокислоты, органические фосфоросодержащие соединения, антибиотики, гуминовые к-ты, витамины, ауксины. Не усваиваются растениями: белки, липиды и др.нерастворимые в воде соединения N, которые входят в состав перегноя. Лучшие источники N-аммиачные соли и соли азотной к-ты.
Аммонификация-процесс разложения белков, аминокислот, мочевины и др соединений, почвенные организмы, осуществляющие этот процесс - аммонификаторы, с их помощью белки разлагаются до аминокислот, которые дезаминируются с образованием NH3. Минерализацию орг.N завершает процесс нитрификации, осуществляемый хемосинтезирующими нитрофицирующими бактериями-аэробами. 2стадии: нитритные бактерии до нитрита, 2ст:нитратные бактерии до нитрата. Корневая система растений способна поглощать аммонийные и нитратные соли. В слабокислой среде(рН5)лучше усваиваются нитраты, в нейтр.среде(рН7)лучше усваиваются аммонийные соли.
Установлено, что высшие растения не способны усваивать молекулярный азот атмосферы. Благодаря деятельности клубеньковых бактерий бобовые растения не только обеспечиваются азотсодержащими соединениями, но и значительно обогащают почву азотом за счет корневых выделений и пожнивных остатков. Высшие растения могут поглощать растворимые азотсодержащие органические соединения — аминокислоты, амиды, короткие полипептиды, но в основном органический азот почвы усваивается после его минерализации.
В водной культуре зеленое растение представляет собой в высшей степени автотрофный организм, способный синтезировать все, в том числе и азотсодержащие, органические соединения. Основными усвояемыми формами азота для высших растений являются ионы аммония и нитрата. Физиологическая особенность процессов усвоения заключается в том, что аммоний сразу после поглощения метаболизируется в корнях, превращаясь в азот аминокислот и амидов. Аммонийная форма азота эффективна только при условии высокой фотосинтетической активности или достаточного количества запасных углеводов. При недостатке органических кислот аммиак не успевает связываться и может быть токсичным для растения. Поступившие нитраты либо запасаются в вакуолях клетках корня, либо подаются с пасокой в надземную систему. Наличие в среде одновременно обеих форм минерального азота приводит к увеличению урожая хлебных злаков. На поглощение растениями нитратов и аммония существенно влияет температура среды. Причем реакция самых разных растений на температурные воздействия оказывается очень сходной: при пониженной температуре относительно или даже абсолютно больше поглощается аммония, чем нитрата, увеличивается метаболическая нагрузка корней по ассимиляции азота
Лекция 6.