

18.Метаболизм кетоновых тел.
Под термином «кетоновые (ацетоновые) тела»
подразумевают ацетоуксусную кислоту (ацетоац
етат) СН3СОСН2СООН, β-оксимасляную кислоту
(β-оксибутират, или D-3-гидроксибутират) СН3СНОНС
Н2СООН и ацетон СН3СОСН3. В печени

Химизм процесса кетогенеза в клетках печени приведен.
• ‒ В реакции (1) осуществляется конденсация 2-х молекул
ацетил-СоА при участии тиолазы. Образуется ацетоацетил
-СоА и удаляется HSCoA.• ‒ На второй стадии происходит в
ключение третьей молекулы ацетил-СоА, но уже при участии
другого фермента ‒ гидроксиметилглутарил-СоА-синтазы
(ГМГ-СоА-синтазы) (2).• ‒ Образующийся -гидрокси--мет
илглутарил-CoA далее расщепляется под действием ГМГ-С
оА-лиазы (3), регенерирующей ацетил-СоА и образующей п
родукт ‒ ацетоацетат.
Кетоновые тела – поставщики «топлива» для мышц, почек
 (в
цикл кребса)
(в
цикл кребса)
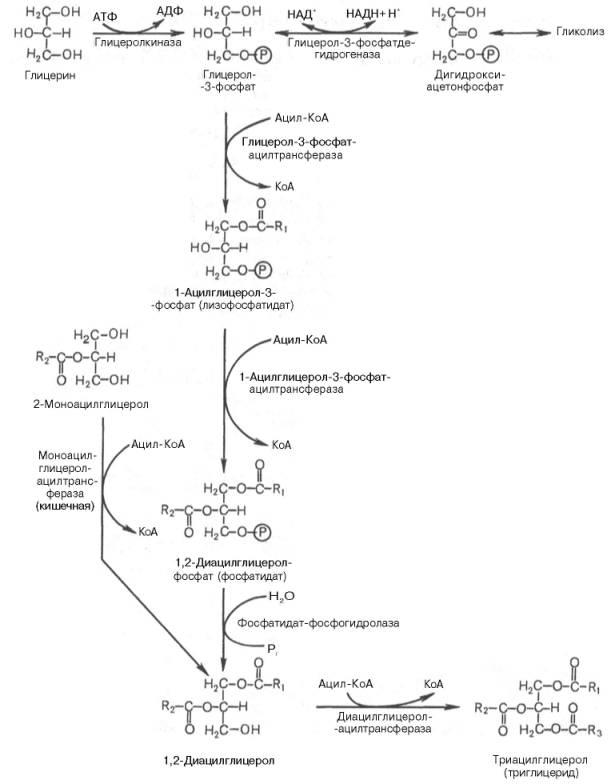
19.Два пути биосинтеза триацилглицеролов.
Синтез триацилглицеролов (липиды) происходит из
глицерола и жирных кислот (преимущественно стеар
иновой, пальмитиновой и олеиновой).
Основными предшественниками для синтеза триацилгли
церолов служат активированные жирные кислоты и глиц
ерол-3-фосфат. Глицерол-3-фосфат образуется либо пр
и прямом фосфорилировании за счет АТР при участии г
лицеролкиназы, либо при восстановлении промежуточн
ого продукта гликолиза ‒ дигидроксиацетон-3-фосфата
ферментом глицерол-3-фосфатдегидрогеназой
20.Биосинтез холестерина. Роль гидроксиметилглутарилСоА
редуктазы в регуляции этого процесса.
Синтез ХС происходит в цитоплазме и микросомах. Биосин тез в печени
составляет приблизительно 80%, в тонком кишечнике синтезируется п
риблизительно 15%, в коже – порядка 5% от количества, продуцируем
ого ежедневно стерола.
Регуляция ключевого фермента синтеза холестерола (ГМГ-Ко
А-редуктазы) происходит разными способами. Фосфорилирова
ние (активн)/дефосфорилирование ГМГ-КоА-редуктазы (неактив)
В результате синтез холестерола в постабсорбтивном периоде и
при голодании ингибируется.
Ингибирование синтеза ГМГ-КоА-редуктазы.Конечный продукт
метаболического пути (холестерол) снижает скорость транскри
пции гена ГМГ-КоА-редуктазы, подавляя таким образом собст
венный синтез. В печени активно идёт синтез жёлчных кислот
из холестерола, поэтому и жёлчные кислоты (как конечные про
дукты синтеза) подавляют активность гена ГМГ-КоА-редуктазы
(рис. 8-67). Так как молекула ГМГ-КоА-редуктазы существует о
коло 3 ч после синтеза, то ингибирование синтеза этого ферме
нта конечным продуктом метаболического пути (холестеролом
) является эффективной регуляцией.
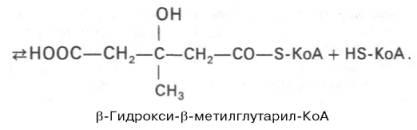
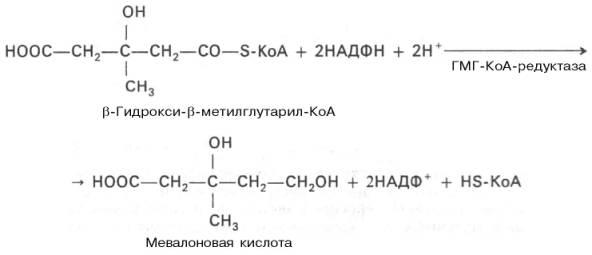
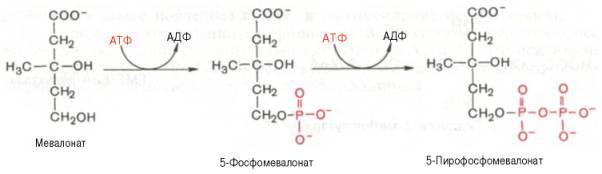
1)включает образование 3-гидрокси-3-метилглутарил-СоА (ГМГ-СоА)
из ацетил-СоА и превращение ГМГ-СоА в мевалонат
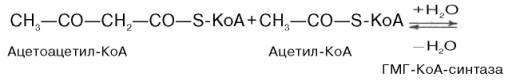
2) превращение мевалоната в фарнезилпирофосфат
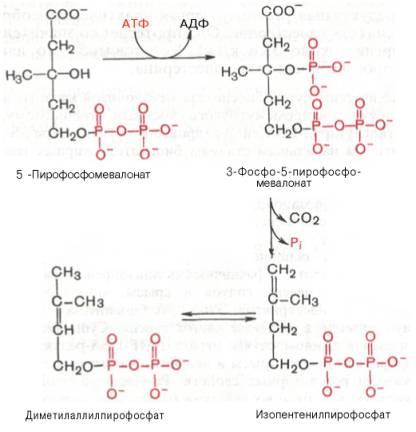
конденсируются с высвобождением пи-рофосфата и образованием
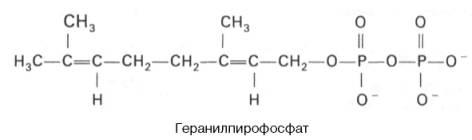
К геранилпирофосфату вновь присоединяется изопентенилпирофосфат.
В результате этой реакции образуется
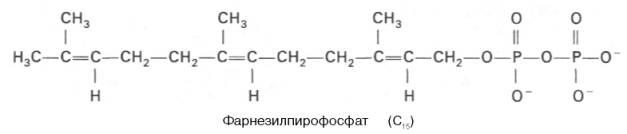
3) образование сквалена из фарнезилпирофосфата и превращение сквалена в ланостерол.
 сквален
под влиянием сквален-оксидоциклазы
сквален
под влиянием сквален-оксидоциклазы
циклизируется с образованием ланостерина.
4)превращение ланостерола в холестерол
Дальнейший процесс превращения ланостерина в холестерин включает
ряд реакций, сопровождающихся удалением трех метильных групп, нас
ыщением двойной связи в боковой цепи и перемещением двойной связи
в кольце В из положения 8, 9 в положение 5, 6 (детально эти последние
еакции еще не изучены
)
