

875
.pdfтакже больше в культурах: 5.52 мг против 5.16 мг. В культурах также присутствует фен гладкой коры у 3.5% крупных деревьев, отсутствующий в природных популяциях.
3.Вероятно, при закладке культур под руководством Ф.А. Теплоухова использовали семена от деревьев с крупными шишками. В потомстве F1 (собственно сами культуры) получено увеличение: длины шишек на 8.2%, массы семени на 6.8% и высоты культур на 1 бонитет. В потомстве F2 (семьи плюс-деревьев) преимущество потомства F2 по высоте составило 4.2%.
4.В популяциях культур обнаружено преимущество матерей, имеющих среднюю массу семени, где лучших семей (высота от 115%) больше в 1.6–1.7 раза. Следовательно, в отношении массы семени в более «европейских» популяциях ели финской действует стабилизирующий отбор.
5.В природных популяциях обнаружено преимущество матерей с малой массой семени, и они дают лучших семей больше в 1.8–2.1 раза. Следовательно,
вболее «сибирских» популяциях ели финской возникает движущий отбор с предпочтением деревьев с легкими семенами.
7.2.Сбег ствола материнского дерева и рост потомства
Внасаждениях складывается разная история их развития. Контрасты в начальной густоте вполне могут привести к формированию ценозов, отличающихся генетически, и их генофонд будет передан следующему поколению. Исследований по этому вопросу у лесных генетиков крайне мало. Пока известны лишь работы А.М. Голикова (2011, 2014).
Воздействие конкуренции в прошлом отражает, а если выразиться образно, регистрирует и «записывает на себя» сбег ствола. Чем выше начальная густота, тем меньше сбег стволов; он тонко реагирует на нее и является индикатором, по которому можно реконструировать историю густоты. Например, зная таксационные показатели и густоту древостоя сейчас и 20 лет назад, можно найти такой же сбег ствола у других древостоев и использовать его для определения их густоты в прошлом. По этому принципу можно восстановить
200
историю развития древостоя с определением его плотности в ранних возрастах и «добраться» до ее истоков даже у спелого древостоя. Именно так подбирались «естественные ряды» при составлении таблиц хода роста с разной начальной густотой по оригинальной методике Г.С. Разина (Рогозин, Разин, 2015). Сбег ствола чрезвычайно важен для характеристики общего состояния древостоя, и в формуле оптимальной густоты выращивания древостоев (Разин, 1989, а.с. на изобретение SU 1464970) сбег ствола использован дважды.
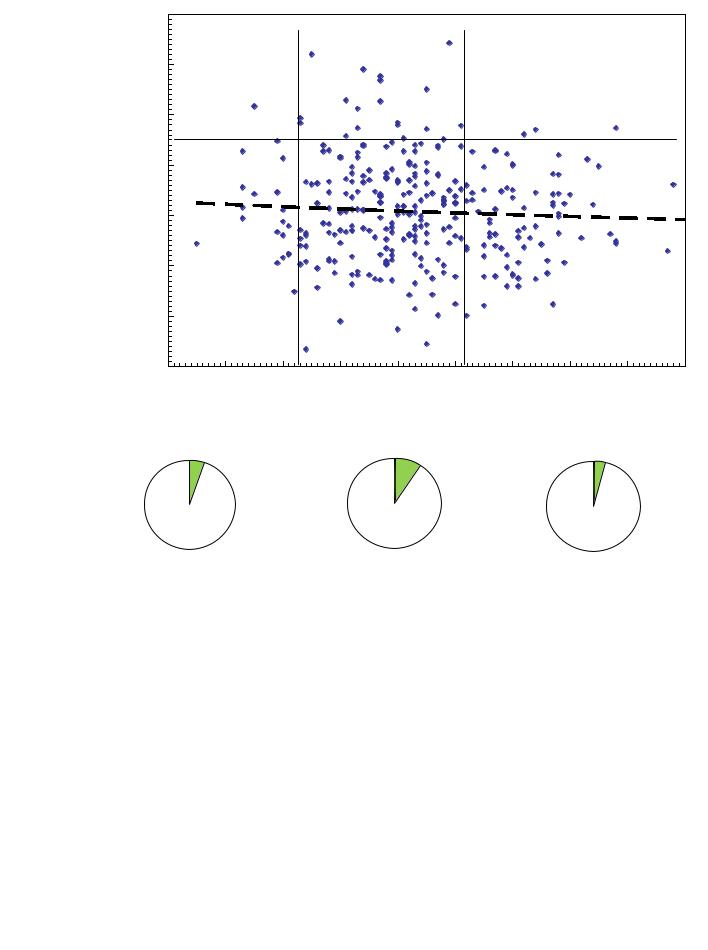
Сбег ствола у плюс-деревьев в естественных популяциях составил в среднем 1.43 см/м против 1.20 см/м в культурах. Различия вызваны как более старшим возрастом деревьев в естественных лесах, который несколько увеличивает сбег, так и большей густотой культур, снижающей его. Между 12 ценопопуляциями средний сбег ствола различался недостоверно, так как объемы выборок оказались недостаточны. Поэтому плюс-деревья объединили в 2 группы: группу из естественных ценозов и группу из культур. Влияние сбега ствола на потомство показано на двух полях корреляций (рис. 7.6 и 7.7).
Результаты весьма интересны. У плюс-деревьев из естественных ценозов в градации среднего сбега (1.22–1.51 см/м) частота лучших семей повышена в 1.9–2.4 раза в сравнению с деревьями с малым (1.0–1.21) и большим (1.52–1.9 см/м) сбегом, при некотором слабом снижении высоты семей от малого сбега к большому, судя по линии тренда (см. рис. 7.6).
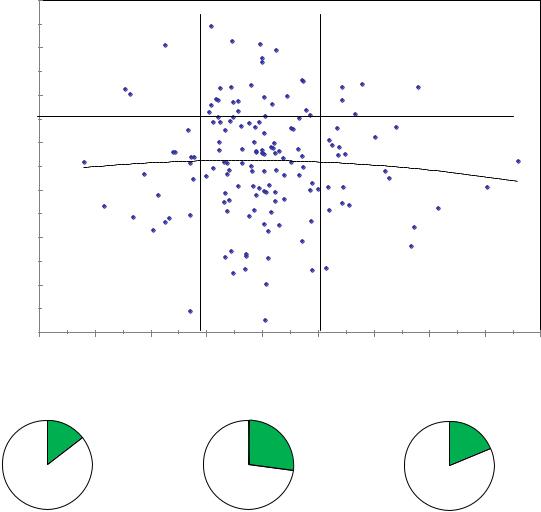
У плюс-деревьев из культур зависимость аналогичная. У матерей со средним сбегом, который в этих, более густых ценозах, значительно меньше (1.1–1.3 см/м) точно также отмечено повышение частоты лучших семей в 1.5– 1.9 раза по сравнению с деревьями с большими или меньшими значениями сбега. Линия тренда здесь показывает практическое отсутствие какого-либо влияния сбега на высоту семей.
Для этих двух совокупностей, однако, имеется и совпадение параметров сбега, в которых наблюдается наилучшее качество потомства: это значения 1.2– 1.3 см/м. Данный параметр можно рекомендовать как общую придержку для отбора плюсовых деревьев ели финской во всех популяциях.
201
|
Эффект этой простой рекомендации весьма значителен: в потомствах |
|||||||||
плюс-деревьев со средним сбегом ствола, имеющего четкие параметры, |
||||||||||
частота лучших семей повышена в среднем в 1.9 раза. |
|
|
|
|||||||
|
140 |
|
|
|
|
|
|
|
|
|
, % |
130 |
|
|
|
|
|
|
|
|
|
потомства |
120 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
летнего |
110 |
|
|
|
|
|
|
|
|
|
|
|
|
|
, |
|
|
|
|
|
|
21- |
100 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
Высота |
90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
|
|
70 |
|
|
|
|
|
|
|
|
|
|
1,0 |
1,1 |
1,2 |
1,3 |
1,4 |
1,5 |
1,6 |
1,7 |
1,8 |
1,9 |
|
|
|
|
Сбег ствола, |
см/м |
|
|
|
|
|
|
|
5 |
|
|
|
9,3 |
|
|
3,9 |
|
|
|
|
|
Доля лучших семей, % |
|
|
|
|
||
Рис. 7.6 – Разделение деревьев из естественных популяций по сбегу ствола на три градации и доля в них лучших семей (с высотами от 115%).
Сбег ствола отражает коллизии конкуренции (в главе 3 мы говорили о том, что он «записывает» историю конкуренции дерева) и характеризует, вероятно, генетические особенности и ценоза, и отдельных деревьев. В условиях высокой густоты и при наблюдаемых малых значениях сбега ствола, например, 1.0–1.2 см/м, в древостое накапливаются родители, толерантные к конкуренции и его генофонд можно назвать «конкурентным».
202
|
140 |
|
|
|
|
|
|
|
|
|
|
130 |
|
|
|
|
|
|
|
|
|
% |
|
|
|
|
|
|
|
|
|
|
потомства, |
120 |
|
|
|
|
|
|
|
|
|
110 |
|
|
|
|
|
|
|
|
|
|
-летнего |
100 |
|
|
|
|
|
|
|
|
|
21 |
|
|
|
|
|
|
|
|
|
|
Высота |
90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
|
|
70 |
|
|
|
|
|
|
|
|
|
|
0,8 |
0,9 |
1,0 |
1,1 |
1,2 |
1,3 |
1,4 |
1,5 |
1,6 |
1,7 |
|
|
|
Сбег ствола матерей, см/м |
|
|
|
|
|||
14,3 |
26,9 |
18,5 |
Доля лучших семей, %
Рис. 7.7 – Разделение деревьев из культур по сбегу ствола на три градации и доля лучших семей (с высотами от 115%) в этих градациях сбега
7.3. Конкурентное давление на матерей и рост их семей
Этот вопрос изучали, проведя картирование соседних деревьев на площадке с радиусом 3.3 м вокруг 79 плюс-деревьев в культурах происхождения «Сепыч-1» и эта работа позволила оценить последствия конкурентного давления на родителей с весьма неожиданной стороны.
Культуры на протяжении 70 лет не подвергались рубкам ухода и выпавшие
203
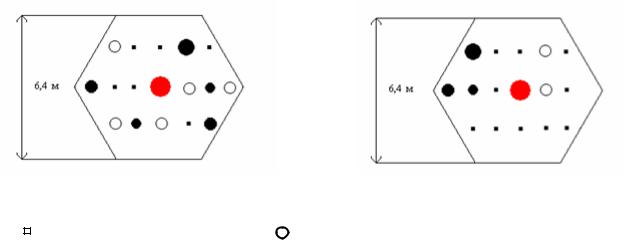
деревья сохранились. Площадки вокруг плюс-деревьев были разделены на две группы по густоте и таксационной полноте. В первой группе со слабой конкуренцией общее количество учтенных стволов на площадке (кроме плюсового дерева) составило в среднем 7.1 шт., а в группе с сильной конкуренцией – 10.9 шт. Как пример, ниже приведены две площадки из этих групп (рис. 7.8).
|
а |
б |
|
посадочные места (пустые); |
погибшие растения и их диаметр; |
|
||
|
 живые растения и их диаметр.
живые растения и их диаметр.
Рис. 7.8 – Примеры сильной (а) и слабой (б) конкуренции вокруг плюсового дерева в культурах 1913 г. на площадке с радиусом 3.3 м
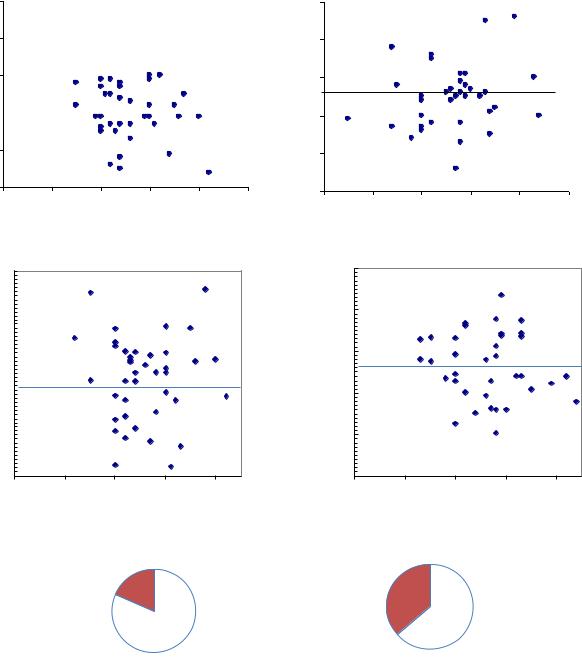
В первой группе с малой густотой и полнотой оказалось 36 плюсовых деревьев, а в группе с большей густотой и полнотой – 43 плюсовых дерева. Уже по 4-летнему потомству было обнаружено преимущество потомства первой группы, которое сохранилось до 21 года в тест-культурах плантационного типа
(+4.6% по средней высоте); при этом доля лучших семей в группе слабой конкуренции оказалась выше в 2 раза: 36.1% против 18.6% (рис. 7.9).
Как видно из проведенного анализа результатов выращивания семей плюсовых деревьев с разной историей конкурентного давления, для роста потомства оказалась решающей не высота матери, а ее формирование с раннего возраста в условиях ослабленной конкуренции, т.е. при отсутствии деревьев в соседних посадочных местах и рядах культур.
204
Сильная конкуренция |
Слабая конкуренция |
Высота потомства, %
Высота потомства , %
130
120
110
100 





90
80 |
|
|
|
|
|
100 |
110 |
120 |
130 |
140 |
150 |
Высота матери, %
130
120
110
100
90
80
100 |
110 |
120 |
130 |
140 |
Высота матери, %
Высота потомства, %
130 |
|
|
|
|
|
120 |
|
|
|
|
|
110 |
|
|
|
|
|
100 |
|
|
|
|
|
90 |
|
|
|
|
|
80 |
|
|
|
|
|
100 |
110 |
120 |
130 |
140 |
150 |
|
|
Высота матери, % |
|
|
|
|
130 |
|
|
|
|
% |
120 |
|
|
|
|
, |
|
|
|
|
|
потомства |
110 |
|
|
|
|
|
|
|
|
|
|
Высота |
100 |
|
|
|
|
90 |
|
|
|
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
100 |
110 |
120 |
130 |
140 |
Высота матери, %
18,6 % |
36.1 % |
Рис. 7.9 – Плюсовые деревья, сформировавшихся в условиях сильной и слабой конкуренции и высота их семей в 4 года (вверху) и в 21 год (внизу), а также доля лучших семей в 21 год; - - - - - средняя высота потомства
Таким образом, сильное конкурентное давление формирует матерей, которые передают потомству пониженную быстроту роста. В то время как при ослабленной конкуренции со стороны соседей рост потомства усиливается, и доля лучших семей в потомстве возрастает практически в 2 раза.
205

7.4. Густота родительских культур и рост их потомства
Родительские культуры, в которых выделялись плюсовые деревья, имели разную историю формирования. Они отличались схемой посадки, приживаемостью, сохранностью и текущей густотой древостоев.
В культурах Сепыч-1, Сепыч-2, Очер, Верещагино посадки проводили по схеме 2.13×1.07 м (1 сажень на 0.5 сажени), а в Нижней Курье – по схеме 2.0×0.6 м с разреживанием в 36 лет до схемы 2.0×1.2 м. При одинаковой схеме посадки ценоз Сепыч-1 сформировался при гораздо большей густоте, так как на этом участке провели дополнения, в отличие от культур Сепыч-2, созданных позднее (в 1916 г.), где такого дополнения уже не было (рис. 7.10).
Рис. 7.10 – Культуры ели Сепыч-1 (слева) сформировались более густыми, так как в них проводили дополнения, в отличие от культур Сепыч-2, где дополнения посадок не было (справа).
Если в родительских культурах присвоить ранги их текущей густоте, то ранги сбега ствола и средних приростов по высоте совпадают с ними полностью (табл. 7.2).
206
Табл. 7.2 – Влияние густоты родительских культур ели на рост их потомства в дочерних испытательных культурах (ИК)
|
|
Родительские культуры |
|
|
|
|
|
Высота в дочерних ИК |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Текущая |
|
|
|
|
|
|
Средний |
редкие ИК: |
густые ИК: |
||||
|
|
|
|
|
Сбег |
|
прирост |
||||||||
|
|
густота |
Средние |
|
|
в ряду 1.0 м |
в ряду 0.7 м |
||||||||
|
|
|
ствола |
по |
|
||||||||||
|
Воз- |
ценоза |
|
|
|
|
условия С2 |
условия В2 |
|||||||
Назва- |
|
|
|
|
|
|
высоте |
||||||||
раст, |
|
|
|
|
|
|
|
|
|
|
|
|
|||
ние |
|
|
|
|
|
|
|
р |
|
р |
|
|
|
|
|
лет |
|
ра |
|
|
|
|
|
|
% |
|
% |
|
|||
|
шт./ |
|
|
|
|
|
а |
см /в |
а |
|
|
||||
|
|
нг |
Н, м |
Д, см |
|
см/м |
|
от конт- |
ранг |
от конт- |
ранг |
||||
|
|
га |
|
|
н |
год |
н |
||||||||
|
|
* |
|
|
|
|
|
роля |
|
роля |
|
||||
|
|
|
|
|
|
|
|
г |
|
г |
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Сепыч-2 |
74 |
770 |
1 |
24,8 |
25,3 |
|
1,020 |
|
1 |
33,5 |
1 |
110,4 |
2 |
99,8 |
5 |
Очер |
88 |
920 |
2 |
25,7 |
25,1 |
|
0,977 |
|
2 |
29,2 |
2 |
110,7 |
1 |
106,2 |
4 |
Сепыч-1 77 1200 3 |
22,2 |
21,1 |
|
0,950 |
|
3 |
28,8 |
3 |
107,8 |
3 |
111,0 |
2 |
|||
Вереща- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
гино |
87 |
1256 |
4 |
24,0 |
21,2 |
|
0,883 |
|
4 |
27,6 |
4 |
97,3 |
5 |
118,9 |
1 |
Нижняя |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Курья |
51 |
1548 |
5 |
18,7 |
16,2 |
|
0,866 |
|
5 |
36,7 |
Н |
100,0 |
4 |
108,1 |
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* - ранги по возрастанию; Н - не ранжировали, так как культуры существенно моложе
Увеличение текущей густоты ценозов сильно повлияло на рост их потомства при выращивании в «редких» испытательных культурах (ИК), снижая высоту культур от 110.4% до 97.3–100%. При этом ранги высот отличаются не более чем на 1 ранг от рангов густоты в родительских культурах.
Иная картина наблюдается в «густых» испытательных культурах. При увеличении густоты родительских популяций 4 потомства из 5 синхронно повышают свои высоты от 99.8 до 118.9%; соответственно, ранги высот и ранги густоты у них также совпадают (см. табл. 7.2, последние графы).
Данные этого сложного опыта представлены также на рисунке (рис. 7.11), из которого становится понятен характер влияния густоты родительских древостоев на рост их потомства в ИК редких (слева) и густых (справа). Средняя высота потомства от трех лучших популяций на левом рисунке с высотами 110.4, 110.7 и 107.4% равна 109.6%, откуда сдвиг при селекции этих популяций по отношению к контролю составит +9.6%.
207

|
120 |
|
|
|
|
|
120 |
|
|
|
118,9 |
|
% |
115 |
|
|
|
|
% |
115 |
|
|
|
|
|
ИК, |
110.4 |
110.7 |
|
|
|
ИК, |
|
|
|
111 |
|
|
|
|
|
|
|
|
|
|
|
||||
редких |
110 |
|
107.8 |
|
|
густых |
110 |
|
|
|
|
108,1 |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
106,2 |
|
|
|
||
105 |
|
|
|
|
105 |
|
|
|
|
|
||
в |
|
|
|
|
в |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||
Высота |
|
|
|
|
100.0 |
Высота |
100 |
99,8 |
|
|
|
|
100 |
|
|
|
|
|
|
|
|
||||
|
|
97.3 |
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
|
95 |
|
|
|
|
|
95 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
770 |
920 |
1200 |
1256 |
1548 |
|
|
770 |
920 |
1200 |
1256 |
1548 |
|
|
|
|
|
|
|
|
|||||
|
Густота родительских популяций, |
шт./га |
|
|
Густота родительских популяций, |
шт./га |
||||||
Рис. 7.11 – Влияние густоты родительских популяций на рост дочерних редких (слева) и густых (справа) испытательных культур (ИК).
Эти три лучшие популяции – самые редкие по их текущей густоте и могут рассматриваться как культуры-аналоги плантационного выращивания ели, и именно их потомство в тест-культурах плантационного типа, где растения размещались 2.5×1.0 м, оказалось самым лучшим. Получается так, что при совпадении конкурентных условий в родительских и дочерних ценозах потомство растет лучше, а при несовпадении – хуже. Условия, отвечающие условиям плантационного выращивания, в родительских ценозах можно либо формировать ранними разреживаниями и ждать, когда плюс-деревья вырастут, либо искать насаждения, сформировавшиеся при минимальной густоте. Распознать такие насаждения-аналоги поможет сбег ствола, который «записывает» историю конкуренции, о чем уже говорилось выше; напомним, что отбор плюс-деревьев по среднему сбегу в 1.5–2.4 раза повышал частоту лучших семей при их выращивании в редких культурах.
Таким образом, наследуется не только быстрота роста, но и память об условиях конкурентной среды, в которой формировался родительский древостой и плюсовые деревья в нем. Можно сказать и так, что как отдельная семья, так и потомство всего насаждения наследуют реакцию родителей о конкурентном давлении: потомство из густых ценопопуляций лучше растет в густых, а потомство из редких – в редких дочерних культурах.
208
7.5.Эпигенетическое влияние родительских популяций и рекомендации для плантационной селекции
Простота и ясность полученных выводов требует, однако, некоторых пояснений. Испытания потомства и их анализ были совсем не простыми.
Прежде всего, дочерние тест-культуры отличались условиями – на основном участке были суглинистые почвы, эдатоп С2, и посадка 2.5×1.0 м (редкие, плантационные культуры) и, казалось бы, совершенно иные условия на страховочном участке, где на супесчаных почвах в эдатопе В2 создавали обычные культуры на вырубке по раскорчеванным полосам шириной 2.0 м. Между полосами было в среднем 9 м, а в полосу растения высаживали тремя рядами по схеме 0.7×0.7 м (рис. 7.12, рис. 7.13).
Различия в росте потомства на этих участках можно было бы объяснить отличиями в типах почвы, если не знать, что решающее влияние оказывает не столько различие в увлажнении, которое для типов С2 и В2 невелико, или в богатстве почвы (которое для С2 и В2 существенно), сколько различие главным образом в плотности дочернего ценоза. Влияние густоты древостоя на его развитие было доказано в главе о законах развития древостоев и, в особенности, при выяснении влияния густоты на численность правых и левых форм, имеющих противоположную адаптацию к густоте и к увлажнению.
Если не знать (или не учитывать) эти новые знания, то да, почвенным условиям будет отведено «главное», а густоте – как бы второе место, т.е. возможно допущение, что «густота, может быть, и влияет, но пока слабо, так как испытательные культуры еще молоды». Влияние почвы настолько широко известно, что ее приоритет даже не обсуждают, а между тем ее влияние надо еще доказать и выявить силу этого влияния конкретными данными. Поясним также, что густая схема посадки на страховочном участке была вынужденной и не была оптимальна для выявления продуктивных семей, но она позволила выяснить реакцию семей на сильную конкуренцию. Причем они испытывали конкуренцию и со стороны лиственных растений, которые местами превышали высоту ели на 1–5 м. Такого на основном участке тест-культур не наблюдалось, и это обстоятельство как раз и было использовано для выяснения эволюции потомства в дочерних ценозах в условиях разного конкурентного давления.
Итоги измерений в испытательных культурах на этих двух участках показаны в таблице (табл. 7.3).
209