

Взаимодействие медиатора с ионотропными рецепторами постсинаптической мембраны
Итак, медиатор диффундирует в щели и связывается с рецепторами постсинаптической мембраны, что приводит к открытию ионных каналов.
В том случае, если увеличивается проницаемость натриевых каналов, то усиливается поступление этих ионов в клетку с последующей деполяризацией возбудимых участков мембраны и возникновением ВПСП (возбуждающий постсинаптический потенциал). Вследствие суммации ВПСП возникает распространяющееся возбуждение в соседнем электровозбудимом участке мембраны. Такие синапсы называют возбуждающими.
В том случае, если увеличивается проницаемость калиевых каналов и каналов для хлора, то наблюдается избыточный выход калия из клетки с одновременным поступлением в нее хлора. Это приводит к гиперполяризации мембраны, снижению ее возбудимости - развитию ТПСП (тормозящий постсинаптический потенциал). Передача нервных импульсов может прекращаться. Такие синапсы называют тормозными.
Исходя из вышеизложенного, можем сказать, что взаимодействие медиатора с рецепторами постсинаптической мембраны может вызывать как возбуждение, так и торможение эффекторной клетки (нейрона).
Инактивирование медиатора - направлено на восстановление постсинаптической мембраны и исходного уровня мп. Оно осуществляется путем:
а) Ферментативного гидролиза с помощью ингибиторов. Например, для АХ - ингибитор холинэстраза, для НА - моноаминоксидаза, катехолоксиметилтрансфераза. Продукты ферментативного гидролиза поступают в кровь или жидкость и циркулируют в них как предшественники.
б) Обратный захват пресинаптическими окончаниями молекул медиатора из синаптической щели и обратный аксонный транспорт - это другой путь удаления медиатора из синаптической щели. НА и адреналин разрушаются ингибиторами в незначительной степени и снова депонируются синаптическими пузырьками в цитоплазме пресинаптических окончаний. Путем обратного аксонного транспорта от окончаний аксона к перикариону движутся крупные пузырьки с «обломками» медиаторов и веществ, подлежащих перевариванию в лизосомах.
Во время инактивирования (полной потери активности) медиатора возбуждение прекращается и возникает снова при наступлении следующей, достаточной для развития ВПСП, порции.
Рецепторы пресинаптической мембраны
- ауторецепторы – на эти рецепторы может действовать сам медиатор, выделяющийся пресинаптическим окончанием;
- гетерорецепторы – на эти рецепторы могут действовать другие медиаторы.
Рецепторы постсинаптической мембраны
Ионотропные рецепторы (прямая синаптическая передача) – это рецепторы, сопряженные с ионными каналами. При этом виде управления рецептор и ионный канал представляют собой единую молекулу. Включают в себя N – ацетилхолиновые рецепторы, рецепторы ГАМК и глицина).
2) Метаботропные рецепторы (непрямая синаптическая передача) - это рецепторы, сопряженные с внутриклеточными ферментативными системами. Когда образуется комплекс медиатор-рецептор, активируется G – белок, а дальше запускается цепочка событий, которые могут приводить или не приводить к открытию или закрытию ионных каналов. Дело в том, что эти рецепторы активируются и медиаторами и гормонами. Они включают в себя М – ацетилхолиновые рецепторы, рецепторы к серотонину, норадреналину, адреналину, гистамину и дофамину.
Электрический синапс = эфапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление пре- и постсинаптических мембран, что обеспечивает прохождение локальных электрических токов. Низкое сопротивление связано с наличием каналов, пересекающих обе мембраны, т.е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами контактирующих мембран, которые соединяются комплементарно. Ток, вызванный пресинаптическим потенциалом действия, раздражает постсинаптическую мембрану, где возникает ВПСП, а затем и потенциал действия. Электрические синапсы формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы). Они обеспечиваю очень быструю передачу возбуждения.
+ Свойства нервных волокон. Распространение возбуждения по безмиелиновым нервным волокнам. Распространение возбуждения по миелиновым волокнам. Законы проведения возбуждения по нервам. Опыт Гассера - Эрлангера.
М еханизмы
проведения возбуждения в безмиелиновых
волокнах. При действии раздражителя
пороговой силы на мембрану безмиелинового
волокна изменяется ее проницаемость
для ионов Nа+, которые мощным потоком
устремляются внутрь волокна. В этом
месте изменяется заряд мембраны
(внутренняя становиться заряженной
положительно, а наружная отрицательно).
Это ведет к возникновению местного
(локального) непрерывного тока от + к –
деполяризует соседний участок. Возникающая
здесь деполяризация пороговая, что
приводит к возникновению ПД.
еханизмы
проведения возбуждения в безмиелиновых
волокнах. При действии раздражителя
пороговой силы на мембрану безмиелинового
волокна изменяется ее проницаемость
для ионов Nа+, которые мощным потоком
устремляются внутрь волокна. В этом
месте изменяется заряд мембраны
(внутренняя становиться заряженной
положительно, а наружная отрицательно).
Это ведет к возникновению местного
(локального) непрерывного тока от + к –
деполяризует соседний участок. Возникающая
здесь деполяризация пороговая, что
приводит к возникновению ПД.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декрементом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
М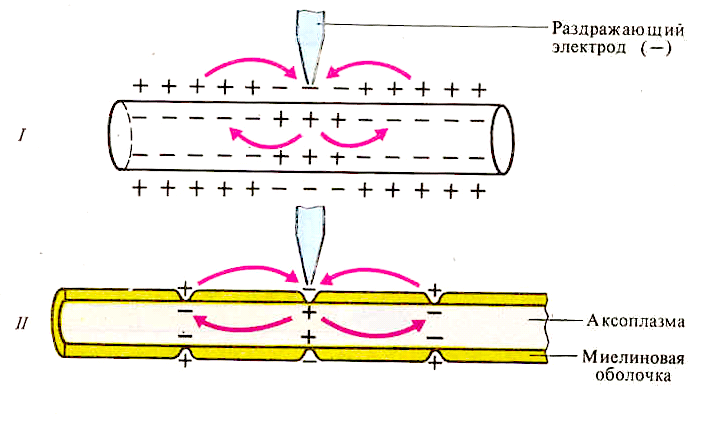 еханизмы
проведения возбуждения в миелиновых
волокнах. В миелинизированном волокне
токи проводятся только в зонах, не
покрытых миелином (перехватах Ранвье).
При действии раздражителя пороговой
силы на мембрану миелинового волокна
в области перехвата Ранвье изменяется
проницаемость для ионов Nа+, которые
мощным потоком устремляются внутрь
волокна. В этом месте изменяется заряд
мембраны, что ведет к возникновению
локального тока. Этот ток идет через
межтканевую жидкость к соседнему
перехвату, где происходит смена заряда.
Таким образом, возбуждение перепрыгивает
с одного участка на другой. Обратное
движение возбуждения невозможно, так
как участок, через который оно прошло,
находится в фазе абсолютной рефрактерности.
еханизмы
проведения возбуждения в миелиновых
волокнах. В миелинизированном волокне
токи проводятся только в зонах, не
покрытых миелином (перехватах Ранвье).
При действии раздражителя пороговой
силы на мембрану миелинового волокна
в области перехвата Ранвье изменяется
проницаемость для ионов Nа+, которые
мощным потоком устремляются внутрь
волокна. В этом месте изменяется заряд
мембраны, что ведет к возникновению
локального тока. Этот ток идет через
межтканевую жидкость к соседнему
перехвату, где происходит смена заряда.
Таким образом, возбуждение перепрыгивает
с одного участка на другой. Обратное
движение возбуждения невозможно, так
как участок, через который оно прошло,
находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно (скачкообразно) от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому.
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декремента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Законы проведения возбуждения по нервам
Закон двустороннего проведения возбуждения по нервному волокну. Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от места его возникновения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т.е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности, проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
В 1934 г. Н. Гассер и Р. Эрлангер провели опыт по анализу составляющих потенциала действия нервного ствола.
Гассер и Эрлангер сделали вывод, что смешанный нерв состоит из трех типов волокон, каждый из которых обладает собственным порогом и скоростью проведения, и вычислили ее для каждого из них. Наиболее быстро проводящие толстые миелиновые волокна отнесены к группе А. Меньшая скорость проведения у более тонких миелиновых волокон группы В. И, наконец, минимальна скорость проведения у тонких безмиелиновых волокон группы С.
+ Функции ЦНС. Координационная деятельность ЦНС. Принципы координации. Интегрирующая роль ЦНС. Нейронный модуль.
Функции ЦНС:
Управление деятельностью опорно-двигательного аппарата. ЦНС регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении восстанавливает ее, инициирует все виды двигательной активности (физическая работа, физкультура, спорт, любое перемещение организма).
Регуляция работы внутренних органов осуществляется вегетативной нервной системой и эндокринными железами; обеспечивает интенсивность их функционирования согласно потребностям организма в различных условиях его жизнедеятельности.
Обеспечение сознания и всех видов психической деятельности. Психическая деятельность - это идеальная, субъективно осознаваемая деятельность организма, осуществляемая с помощью нейрофизиологических процессов. И. П. Павлов ввел представление о высшей и низшей нервной деятельности. Высшая нервная деятельность — это совокупность нейрофизиологических процессов, обеспечивающих сознание, подсознательную переработку информации и целенаправленное поведение организма в окружающей среде. Психическая деятельность осуществляется с помощью высшей нервной деятельности и протекает осознанно, т.е. во время бодрствования, независимо от того, сопровождается она физической работой или нет. Высшая нервная деятельность протекает во время бодрствования и сна. Низшая нервная деятельность - это совокупность нейрофизиологических процессов, обеспечивающих осуществление безусловных рефлексов.
Формирование взаимодействия организма с окружающей средой. Это реализуется, например, с помощью избегания или избавления от неприятных раздражителей (защитные реакции организма), регуляции интенсивности обмена веществ при изменении температуры окружающей среды. Изменения внутренней среды организма, воспринимаемые субъективно в виде ощущений, также побуждают организм к той или иной целенаправленной двигательной активности. Так, например, в случае недостатка воды и при повышении осмотического давления жидкостей организма возникает жажда, которая инициирует поведение, направленное на поиск и прием воды. Любая деятельность самой ЦНС реализуется в конечном итоге с помощью функционирования отдельных клеток.
Координационная деятельность – это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Основа координационной деятельности (КД) – взаимодействие процессов возбуждения и торможения. Функции КД:
1) обеспечивает четкое выполнение определенных функций, рефлексов;
2) обеспечивает последовательное включение в работу различных нервных центров для обеспечения сложных форм деятельности;
3) обеспечивает согласованную работу различных нервных центров (при акте глотания в момент глотания задерживается дыхание, при возбуждении центра глотания тормозится центр дыхания).
Принципы координационной деятельности:
1) Факторы структурно-функциональной связи
а) Прямая связь – управление другим центром или рабочим органом с помощью посылки к ним эфферентных импульсов (команд);
б) Обратная связь - управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них;
в) Реципрокная связь – связь, обеспечивающая торможение центра – антагониста при возбуждении центра;
г) Принципы модульной (ансамблевой) структурно-функциональной организации ЦНС. Нейронный ансамбль (модуль) – совокупность повторяющихся локальных нейронных сетей, образовывающих и передающих информацию с помощью внешних и внутренних связей.
2) Фактор субординации; Основная тенденция в эволюции нервной системы проявляется в сосредоточении функций регуляции и координации в высших отделах ЦНС – цефализация функций нервной системы
3) Фактор силы; Импульсы, приходящие в ЦНС по разным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным или эфферентным нейронам. Один и тот же мотонейрон может возбуждаться импульсами, приходящими от различных рецепторов (зрительных, слуховых, тактильных), т.е. участвовать во многих рефлекторных реакциях (включаться в различные рефлекторные дуги). Так, например, мотонейроны, иннервирующие дыхательную мускулатуру, помимо обеспечения вдоха участвуют в таких рефлекорных реакциях, как чихание, кашель и др. причем рефлекс будет идти о наиболее сильного возбудителя.
4) Односторонне проведение возбуждения;
5) Феномен облегчения; возбуждение быстрее проходит по протоптанным путям.
6) Доминанта – обеспечивает автоматизированное выполнение двигательных актов пригимнастических элементов, в процессе трудовой деятельности человека. Был открыт А.А. Ухтомским, который обнаружил, что раздражение афферентного нерва (или коркового центра), ведущего к сокращению мышц конечностей при переполнении у животного кишечника, приводит к акту дефекации. В данной ситуации рефлекторное возбуждение центра дефекации подавляет, тормозит двигательные центры, а центр дефекации начинает реагировать на посторонние для него сигналы. К доминантному очагу конвергируют возбуждения из различных областей ЦНС, а способность других центров реагировать на сигналы, приходящие к ним, затормаживается. Благодаря этому создаются условия для формирования определенной реакции организма на раздражитель, имеющий наибольшее биологическое значение, т.е. удовлетворяющий жизненно важную потребность.
Интегрирующая роль нервной системы – это соподчинение и объединение тканей и органов в центрально-периферическую систему, деятельность которой направлена на достижение полезного для организма приспособительного результата.
Уровни ЦНС:
Нейрон;
Нейрональный ансамбль;
Нервный центр;
Высший уровень интеграции.
+ Нейрон как структурно-функциональная единица ЦНС. Функции нейрона. Виды нейронов. Нейроглия и ее функции. Методы исследования функций ЦНС.
Нейрон как структурно-функциональная единица ЦНС
Структура, обеспечивающая синтез макромолекул, которые транспортируются по аксону и дендритам – сома (тело нейрона), выполняющая трофическую функцию;
Структуры, воспринимающие импульсы от других нервных клеток – тело и дендриты нейрона с расположенными на них шипиками;
Структура, где возникает ПД – аксонный холмик;
Структура, проводящая возбуждение к другому нейрону – аксон;
Структуры, передающие импульсы на другие клетки – синапсы.
Функции нейрона. Хранение, интеграция, воспроизведение информации; синтез БАВ; генерация электрических импульсов; проведение возбуждения; аксонный транспорт; выделение медиатора (6).
Виды нейронов:
Афферентные – анализируют сигналы, идущие с рецепторов;
Эфферентные – дают команду эффектору;
Вставочные - осуществляют передачу сигнала от одного нейрона к другому, способствуют распределению сигналов по нейронным сетям, осуществляют торможение и принятие решения, поддерживают активность отдельных нервных центров;
Морфологически:
биполярные(один аксон, один дендрит);
униполярные(один отросток-аксон);
псевдоуниполятные(один отросток Т-образно раздвоенный при выходе из клетки);
мультиполярные(один аксон и много дентритов).
Н ейроглия:
- совокупность всех клеточных элементов
нервной ткани, кроме нейронов.
ейроглия:
- совокупность всех клеточных элементов
нервной ткани, кроме нейронов.
Методы исследования функций ЦНС
1) Метод разрушения. Используя этот метод, можно установить какие функции ЦНС выпадают после оперативного вмешательства и какие сохраняются. Однако разрушение и экстирпация являются грубыми вмешательствами, и они сопровождаются существенными изменениями функций ЦНС и организма в целом.
2) Метод перерезки — дает возможность изучить значение в деятельности того или иного отдела ЦНС, влияний, поступающих от других ее отделов. Полная перерезка, например, спинного мозга или ствола мозга разобщает вышележащие отделы ЦНС от нижележащих и позволяет изучить рефлекторные реакции, которые осуществляются нервными центрами, расположенными ниже места перерезки. Перерезка и локальное повреждение отдельных нервных центров производится не только в условиях эксперимента, но и в нейрохирургической клинике в качестве лечебных мероприятий.
3) Метод раздражения позволяет изучить функциональное значение различных образований ЦНС. При раздражении (химическом, электрическом и т.д.) определенных структур мозга можно наблюдать возникновение, особенности проявления и характер распространения процессов возбуждения.
4) Электрографические методы. К этим методам исследования функций ЦНС относятся:
А) электроэнцефалография — метод регистрации суммарной электрической активности различных отделов головного мозга.
Б) Метод локального отведения потенциалов, когда биотоки регистрируются с определенных ядерных образований либо в остром эксперименте, либо после предварительного вживления электродов – в хроническом опыте.
В) Метод вызванных потенциалов, когда регистрируется электрическая активность определенных структур мозга при стимуляции рецепторов, нервов, подкорковых структур. Различают первичные (ПО) и поздние или вторичные (ВО) вызванные потенциалы.
5) Исследование рефлексов (например, коленный, ахиллов, брюшные и т.д).
6)Фармакологические методы с использованием нейроактивных веществ медиаторной или пептидной природы, гормонов и лекарственных веществ, обладающих специфическим влиянием на рецепторы (например, миметики — адрено-,- холино- или блокаторы этих рецепторов) ЦНС.
7) Биохимические методы.
+ Современное представление о механизмах центрального торможения. Классификация видов торможения в ЦНС. Постсинаптическое торможение в ЦНС. Пресинаптическое торможение. Возвратное и реципрокное торможение.
Торможение в ЦНС – активный нервный процесс, результатом которого является прекращение или ослабление возбуждения (Сеченов, 1863).
Гольц (1870) – обнаружил проявление торможения у спинальной лягушки.
Мегун (1944) установил, что раздражения медиальной части РФ продолговатого мозга тормозит рефлекторную активность спинного мозга.
Классификация торможения в ЦНС:
По электрическому состоянию мембраны
Гиперполяризационное. Увеличение проницаемости для К и СЛ.
Деполяризационное. Инактивация На-каналов.
По отношению к синапсу
Постсинаптическое – основной вид торможения, заключается в том, что в нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону импульса выделяется медиатор, который гиперполяризует постсинаптическую мембрану другого, возбуждающего нейрона. В мембране последнего деполяризация не может достигнуть критического уровня и поэтому возбуждение по нейрону не распространяется. Открыл Д.Экклс при регистрации потенциалов мотонейронов спинного мозга кошки во время раздражения мышечных афферентов Iа (мышц-разгибателей).
Механизм постсинаптического торможения.
Возбудимость клетки от ТПСП уменьшается, потому что увеличивается пороговый потенциал, т.к. КУД остается на прежнем уровне, а МП возрастает.
В спинном мозге глицин вырабатывается клетками Реншоу в синапсах. Действуя на рецептор постсинаптической мембраны. Глицин увеличивает ее проницаемость для Clˉ, при этом Clˉ поступает в клетку согласно концентрационному градиенту, в результате чего и развивается гиперполяризация.
 При
действии ГАМК на постсинаптическую
мембрану ТПСП развивается в результате
входа Clˉ в клетку
или выхода К⁺ из клетки.
При
действии ГАМК на постсинаптическую
мембрану ТПСП развивается в результате
входа Clˉ в клетку
или выхода К⁺ из клетки.
Виды постсинаптического торможения:
В
 озвратное
– это торможение, при котором тормозные
вставочные нейроны действуют на те же
нервные клетки, которые их активируют.
Торможение тем глубже, чем сильнее было
предшествующие возбуждение. Возвратное
торможение осуществляется вставочными
тормозными клетками (клетками Реншоу).
Аксоны мотонейронов часто дают
коллатерали (ответвления), оканчивающиеся
на клетках Реншоу. Аксоны клеток Реншоу
оканчиваются на теле или дендритах
этого мотонейрона, образуя тормозные
синапсы. Возбуждение, возникающее в
мотонейроне, распространяется по
прямому пути к скелетной мускулатуре,
а также по коллатералям к тормозящему
нейрону, которые посылают импульсы к
мотонейронам и тормозят их. Чем сильнее
возбуждение мотонейрона, тем сильнее
возбуждаются клетки Реншоу и тем более
интенсивно они оказывают свое тормозящее
действие, что предохраняет нервные
клетки от перевозбуждения.
озвратное
– это торможение, при котором тормозные
вставочные нейроны действуют на те же
нервные клетки, которые их активируют.
Торможение тем глубже, чем сильнее было
предшествующие возбуждение. Возвратное
торможение осуществляется вставочными
тормозными клетками (клетками Реншоу).
Аксоны мотонейронов часто дают
коллатерали (ответвления), оканчивающиеся
на клетках Реншоу. Аксоны клеток Реншоу
оканчиваются на теле или дендритах
этого мотонейрона, образуя тормозные
синапсы. Возбуждение, возникающее в
мотонейроне, распространяется по
прямому пути к скелетной мускулатуре,
а также по коллатералям к тормозящему
нейрону, которые посылают импульсы к
мотонейронам и тормозят их. Чем сильнее
возбуждение мотонейрона, тем сильнее
возбуждаются клетки Реншоу и тем более
интенсивно они оказывают свое тормозящее
действие, что предохраняет нервные
клетки от перевозбуждения.Л
 атеральное
– это тормозные вставочные нейроны
соединены так, что они активируются
импульсами от возбужденного центра и
влияют на соседние клетки с такими же
функциями. Называются так потому что,
образующаяся зона торможения располагается
«сбоку» по отношению к возбужденному
нейрону и инициируется им.
атеральное
– это тормозные вставочные нейроны
соединены так, что они активируются
импульсами от возбужденного центра и
влияют на соседние клетки с такими же
функциями. Называются так потому что,
образующаяся зона торможения располагается
«сбоку» по отношению к возбужденному
нейрону и инициируется им.Реципрокное (прямое) вызывает угнетение центра-антагониста. Возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку Реншоу – к центру мышцы-антагониста – разгибателю, что предотвращает её сокращение. Например, ходьба.
Параллельное - когда возбуждение блокирует само себя, за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону !!! то есть оно блокирует ту клетку, к которой идет торможение!!!, который активировался этим же возбуждением.
В реальной действительности вариантов торможения больше, что определяется множеством связей различных нейронов.
Пресинаптическое. Выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов.
Опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых α-мотонейронов. МП и возбудимость исследуемых мотонейронов не изменялись, либо регистрировался низкоамплитудный ВПСП, т.к. в опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги, было очевидно: они возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что и определяет название этого вида. Возможно паралелльное и латеральное торможение.
Все варианты пост- и пресинаптического торможения объединяют в 2 группы:
1) Когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное, возвратное);
2 )
Когда блокируются другие нервные
элементы под влиянием импульсов от
соседних возбуждающих нейронов с
включением тормозных клеток (латеральное,
прямое) т.к. тормозные клетки сами
могут быть заторможены другими тормозными
нейронами, это может облегчить
распространение возбуждения.
)
Когда блокируются другие нервные
элементы под влиянием импульсов от
соседних возбуждающих нейронов с
включением тормозных клеток (латеральное,
прямое) т.к. тормозные клетки сами
могут быть заторможены другими тормозными
нейронами, это может облегчить
распространение возбуждения.
по нейрональной организации:
Поступательное торможение обусловлено тем, что на пути следования возбуждения встречается тормозной нейрон.
Возвратное торможение.
Латеральное торможение.
Роль торможения:
1) Оба вида торможения со всеми видами их разновидностями выполняют охранительную роль (отсутствие привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС);
2)Играет важную роль в обработке поступающей в ЦНС информации;
3)Обеспечение координационной деятельности ЦНС.
+ Организация нервной системы. Свойства нервных центров. Особенности нервного волокна. Классификация рефлексов. Рефлекторная дуга.
Организация нервной системы. Нервная система подразделяется на две части: центральную и периферическую. ЦНС состоит из нейронов, их отростков и глии, расположенных в головном и спинном мозге. Периферическая нервная система образована нейронами, их отростками и глией, находящимися за пределами ЦНС.
Свойства нервных центров.
Иррадиация возбуждения – распространение потенциала действия в нервных центрах посредством вставочных нейронов.
Суммация возбуждения – способность нервного центра отвечать на действие нескольких афферентных стимулов, каждый из которых в отдельности не может вызвать рефлекторного ответа.
Трансформация ритма возбуждения – изменение числа потенциалов действия, возникающих в нейронах центра на выходе, относительно числа потенциалов действия, поступающих на вход данного центра. Непосредственным местом трансформации ритма и частоты импульсов являются синапсы, через которые передается возбуждение.
Окклюзия – уменьшение эффекта одновременного действия двух сильных афферентных возбуждающих входов в нервный центр по сравнению с суммой их раздельных эффектов. Окклюзия наблюдается при одновременном раздражении нейронов, входящих в центральные зоны разных нейронных популяций данного нервного центра.
Пространственное облегчение – это усиление эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в нервный центр по сравнению с суммой их раздельных эффектов. Облегчение связано с активацией нейронов, входящих в периферическую «кайму» разных нейронов данного центра.
Посттетаническая потенциация – усиление рефлекторного ответа под влиянием предшествующих частых ритмических (т. н. тетанизирующих) раздражений. Механизм заключается в облегчении синаптического проведения возбуждения вследствие увеличения количества квантов медиатора, высвобождаемых пресинаптическим нервным окончанием после предшествующего ритмического раздражения. Последействие – продолжение ответной рефлекторной реакции в течение некоторого времени после прекращения действия раздражителя.
Утомляемость – постепенное снижение интенсивности рефлекторного ответа при продолжительном раздражении афферентных входов нервного центра. Данное свойство связано с расходом энергетических ресурсов и нейромедиаторов в синаптических окончаниях нейронов в нервном центре.
Избирательная чувствительность к действию физиологически активных веществ (токсинов, наркотических веществ, лекарств, нейромедиаторов, нейропептидов, цитокинов и др.).
Высокая чувствительность к гипоксии (различается у нервных центров спинного мозга, мозгового ствола и коры больших полушарий). Наименьший временной предел реанимации после аноксии характерен для коры больших полушарий (5–10 мин).
Рефлекс – это ответная реакция организма на раздражение рецепторов, осуществляемая при участии ЦНС.
Особенности нервного волокна.
группа А (альфа, бета, гамма, дельта) - самый короткий потенциал действия, самая толстая миелиновая оболочка, самая высокая скорость проведения возбуждения;
группа В - миелиновая оболочка менее выражена;
группа С - без миелиновой оболочки.
По ряду признаков рефлексы могут быть разделены на группы
По типу образования: условные и безусловные рефлексы
По видам рецепторов: экстероцептивные (кожные, зрительные, слуховые, обонятельные), интероцептивные (с рецепторов внутренних органов) и проприоцептивные (с рецепторов мышц, сухожилий, суставов)
По эффекторам: соматические, или двигательные (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; вегетативные внутренних органов — пищеварительные, сердечнососудистые, выделительные, секреторные и др.
По биологической значимости: оборонительные, или защитные, пищеварительные, половые, ориентировочные.
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный).
По характеру влияний на деятельность эффектора: возбудительные — вызывающими и усиливающими (облегчающими) его деятельность, тормозные — ослабляющими и подавляющими её (например, рефлекторное учащение сердечного ритма симпатическим нервом и урежение его или остановка сердца — блуждающим).
По анатомическому расположению центральной части рефлекторных дуг различают спинальные рефлексы и рефлексы головного мозга. В осуществлении спинальных рефлексов участвуют нейроны, расположенные в спинном мозге. Пример простейшего спинального рефлекса — отдергивание руки от острой булавки. Рефлексы головного мозга осуществляются при участии нейронов головного мозга. Среди них различают бульбарные, осуществляемые при участии нейронов продолговатого мозга; мезэнцефальные — с участием нейронов среднего мозга; кортикальные — с участием нейронов коры больших полушарий головного мозга.
Рефлекторная дуга (нервная дуга) — путь, проходимый нервными импульсами при осуществлении рефлекса.
Рефлекторная дуга состоит из шести компонентов: рецепторов, афферентного пути, рефлекторного центра, эфферентного пути, эффектора (рабочего органа), обратной связи.
Рефлекторные дуги могут быть двух видов:
1) простые – моносинаптические рефлекторные дуги (рефлекторная дуга сухожильного рефлекса), состоящие из 2 нейронов (рецепторного (афферентного) и эффекторного), между ними имеется 1 синапс;
2) сложные – полисинаптические рефлекторные дуги. В их состав входят 3 нейрона (их может быть и больше) – рецепторный, один или несколько вставочных и эффекторный.
+ Первые сведения о структуре и функциях вегетативной нервной системы. Отличие ВНС от соматической. Адаптационно-трофическая функция нервной системы. Эрготрофная функция симпатической нервной системы. Трофотропная функция парасимпатической нервной системы.
ВНС - часть общей нервной системы, имеющая отношение преимущественно к регуляции вегетативных функций: сокращению гладких мышц, секреции железистого аппарата, функционированию соединительной ткани внутренних органов, тонусу сосудов.
Гален – первые сведения о структуре и функции ВНС;
Рейл – ввел понятие «ВНС»;
Ленгли – применил никотиновый метод, ввел понятия «пре- и постганглионарные нервные волокна», разделил ВНС на 2 отдела.
Отличие ВНС от соматической
Влияние ВНС на организм не находится под непосредственным контролем сознания.
Регуляция функций внутренних органов ВНС может осуществляться при полном нарушении связи с ЦНС, т.к. эффекторный нейрон ВНС находится за пределами ЦНС.
Генерализованный характер распространения возбуждения в периферическом отделе ВНС.
Низкая скорость проведения возбуждения в вегетативных нервах 3-14 м/с (соматических – 120 м/с).
Низкая лабильность нейронов вегетативных ганглиев - 10-15 имп/с (у соматической – 200 имп/с).
6) В ВНС эффекторный нейрон располагается за пределами головного и спинного мозга и находится в ганглиях (паравертебрально, превертебрально, интрамурально), а в соматических - в ЦНС (серое вещество спинного мозга);
7) Афферентное звено дуги вегетативного рефлекса может быть образовано как собственными вегетативными, так и соматическими афферентами (в дуге соматического - только собственные).
8) В дуге вегетативного рефлекса слабее выражена сегментированность.
Адаптационно-трофическая функция симпатической нервной системы. Феномер Обрели-Гинецинского связан с значением симпатической иннервации для скелетных мышц. Содержит 2 компонента:
Адаптационный – приспособление органов к нагрузкам, оказывает на органы трофическое действие, изменение скорости протекания метаболических процессов.
Трофический. Если на 100% изнеможённую мышцу подействовать СНС, а затем через моторные нервы, то мышца будет сокращаться, тк снс стимулирует потребление О2, чувствительность к АХ, укорочение хронаксии.
Эрготрофная функция симпатической нервной системы. Способствует приспособлению к меняющимся условиям внешней среды (голоду, холоду), обеспечивает гомеостаз в период активной физической и психической деятельности. Она запускает процессы катаболизма (процессы расщепления ресурсов). Степень этой комплексной реакции зависит от важности, значимости новизны ситуации, с которой встретился организм. Эрготропная система сначала активизируется невральной фазой - с помощью симпатической нервной системы, в том числе выбрасывающей адреналин. Затем ее активность усиливается вторичной гуморальной фазой (выделением гормонов), зависящей от уровня циркулирующего адреналина.
Трофотропная функция парасимпатической нервной системы направлена на поддержание динамического постоянства внутренней среды организма (его физико-химических, биохимических, ферментативных, гуморальных и других констант).
+ Понятия вегетативной и соматической нервной системы. Структурно-функциональные особенности ВНС. Структурно-функциональные особенности соматической нервной системы. Рефлекторная дуга вегетативного рефлекса. Рефлекторная дуга соматического рефлекса.
Понятия вегетативной и соматической нервной системы. Соматическая нервная система регулирует работу поперечнополосатой мышечной ткани скелетных мышц. Высшим центром соматической нервной системы является кора больших полушарий. Сюда стекается вся информация от органов чувств и внутренней среде организма. Здесь изыскиваются способы удовлетворения потребностей. В лобных долях коры созревает план будущих действий, который реализуется соматической нервной системой. Цели человека много сложнее, чем цели животных, но и они в конечном счёте сводятся к мышечному движению — будь то работа на станке, письмо, речевое общение или даже чтение (движение глаз, произнесение слов про себя и т. д.). Приспособление к природной и социальной среде, связанное с изменением поведения, осуществляется соматической нервной системой.
А втономная
(вегетативная) нервная система, как и
соматическая, имеет центральную и
периферическую части. Высшим органом
автономной нервной системы считается
гипоталамус. Он регулирует не только
автономную нервную систему, но и
эндокринные железы через гипофиз.
Автономная нервная система подразделяется
на два подотдела — симпатический и
парасимпатический.
втономная
(вегетативная) нервная система, как и
соматическая, имеет центральную и
периферическую части. Высшим органом
автономной нервной системы считается
гипоталамус. Он регулирует не только
автономную нервную систему, но и
эндокринные железы через гипофиз.
Автономная нервная система подразделяется
на два подотдела — симпатический и
парасимпатический.
Рефлекторная дуга вегетативного нервного рефлекса.
Чувствительные нейроны представлены псевдоуниполярными клетками спинномозгового узла, так же как и в соматической нервной системе. Их периферические отростки заканчиваются рецепторами в органах. Центральный отросток чувствительного нейрона в составе заднего корешка вступает в спинной мозг и нервный импульс переключается на вставочный нейрон, клеточное тело которого расположено в боковых рогах (латерально-промежуточное ядро тораколюмбального или сакрального отделов) серого вещества спинного мозга.
Вставочный нейрон отдает аксон, который покидает спинной мозг в составе передних корешков и достигает одного из вегетативных узлов, где вступает в контакт с эффекторным (двигательным) нейроном. Таким образом, второе звено вегетативной рефлекторной дуги отличается от соматической, во-первых, местом локализации тела вставочного нейрона, во-вторых, протяженностью и положением аксона, который в отличие от соматической нервной системы никогда не остается в пределах спинного мозга.
Еще большие различия в строении третьего звена рефлекторной дуги. В отличие от соматической рефлекторной дуги, где двигательные нейроны расположены в передних рогах спинного мозга, для вегетативной рефлекторной дуги характерно расположение двигательного нейрона за пределами центральной нервной системы - в вегетативных узлах, аксоны которых направляются к рабочему органу, а это значит, что весь эфферентный путь подразделяется на два участка: предузловой (преганглионарный) - аксон вставочного нейрона и послеузловой (постганглионарный) - аксон двигательного нейрона вегетативного узла. Таким образом, в вегетативной рефлекторной дуге эфферентный периферический путь двухнейронный.
В простой трехнейронной вегетативной рефлекторной дуге, замыкающейся в пределах мозгового ствола, тело первого нейрона располагается в чувствительных узлах черепных нервов, второго - в вегетативных ядрах черепных нервов (мезенцефалический и бульбарный отделы) и третьего - в вегетативных узлах.
Рефлекторная дуга соматической нервной системы.

+ Физиологические свойства симпатического отдела ВНС. Физиологические свойства парасимпатического отдела. Роль метасимпатического отдела. Влияние симпатического, парасимпатического и метасимпатического отделов на иннервируемые органы. Синергизм и относительный антагонизм, их влияние.
Физиологические свойства СНС:
Охватывают несколько органов, генерализованные влияния за счет разветвления аксонов.
Симпатическая нервная система оказывает преимущественно активирующие влияния на функции иннервируемых органов.
Симпатическая нервная система усиливает катаболические реакции организма.
Симпатическая нервная система участвует в формировании таких целостных состояний, как агрессия, стресс, болевые реакции. Влияние симпатической нервной системы мобилизует организм на борьбу и бегство, активное взаимодействие с окружающим миром.
В симпатической нервной системе передача с пре- на ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — посредством норадреналина. Исключение составляют постганглионарные симпатические волокна к потовым железам, в окончаниях которых выделяется ацетилхолин.
Симпатическая нервная система представлена на периферии на значительном протяжении постганглионарными волокнами типа С, вследствие этого возбуждение по симпатической нервной системе распространяется относительно медленнее, чем по парасимпатической нервной системе.
Эффекты симпатической нервной системы по сравнению с парасимпатической более продолжительны.
Электрические потенциалы в симпатических ганглиях характеризуются продолжительными следовыми явлениями. При стимуляции преганглионарных волокон в симпатических ганглиях сначала регистрируется быстрый возбуждающий постсинаптический потенциал (ВПСП), который сменяется пролонгированным медленным ТПСП. Вслед за этим снова регистрируется медленный возбуждающий постсинаптический потенциал. Медленный ВПСП продолжается минуты. Установлено, что начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы. Медленный ТПСП определяется действием дофамина, который секретируется малыми по размеру интернейронами ганглия. Интернейроны, в свою очередь, активируются через мускариновые рецепторы.
Физиологические свойства парасимпатической нервной системы:
В отличие от симпатической нервной системы мультипликация парасимпатических возбуждений наблюдается в районе иннервируемого органа, поэтому это влияние направленное и локальное.
Парасимпатическая нервная система оказывает успокаивающее, расслабляющее действие на функции организма.
Полагают, что парасимпатическая нервная система усиливает преимущественно анаболические реакции.
В парасимпатической системе медиатором в преганглионарных и постганглионарных волокнах служит ацетилхолин.
Парасимпатическая нервная система представлена на значительном расстоянии преганглионарными волокнами типа Б. Вследствие этого возбуждения по парасимпатической нервной системе распространяются значительно быстрее, чем по симпатической нервной системе.
Эффекты парасимпатической нервной системы по сравнению с симпатической менее продолжительны.
Генерализованные гуморальные влияния вегетативной нервной системы. Выделяющиеся при возбуждении вегетативной нервной системы медиаторы оказывают свое действие не только на постсинаптические мембраны, где они быстро разрушаются соответствующими ферментами, но и могут, особенно при блокаде этих ферментов, поступать в кровь, оказывая генерализованное влияние на многие органы, имеющие к ним соответствующие рецепторы.
Метасимпатичсекая НС. Сплетения Ауэрбаха и Мейнснера. Расположена в слоях полых органов, обладает автоматией. Преганглионарные волокна выделяют ацетилхолин и норадреналин, постганглионарные – АТФ и аденозин, ацетилхолин, норадреналин, серотонин, дофамин, адреналин, гистамин и т.д. Главная роль принадлежит АТФ и аденозину. Такая регуляция является базовой, в то же время в случае необходимости она может корректироваться более высокими уровнями ВНС. Рефлекторные дуги полностью находятся в органе и не имеют выхода из ЦНС. Т. е. МНС - это комплекс микроганглионарных образований, расположенных в стенках внутренних органов, обладающих моторной активностью.
Типы нейронов в МНС по Догелю:
Нейроны 1 типа - эфферентные нейроны, аксон которых непосредственно контактирует с мышечной клеткой;
Нейроны 2 типа – афферентные нейроны, аксоны которых могут переключаться на нейроны 1 типа, либо аксон может идти к пара – и превертебральному ганглиям, переключаясь на другие нейроны, либо могут доходить до спинного мозга и переключаться на другие нейроны.
Нейроны 3 типа – ассоциативные нейроны.
Функции и роль:
Осуществляет передачу центральных влияний за счет того, что парасимпатические и симпатические волокна могут контактировать с метасимпатикой и коррегировать ее влияние на объекты управления;
Может выполнять роль самостоятельного интегрирующего образования, т.к. в ней имеются готовые рефлекторные дуги.
Р ОЛЬ
– осуществление механизмов, обеспечивающих
относительное динамическое постоянство
внутренней среды и устойчивость основных
физиологических функций.
ОЛЬ
– осуществление механизмов, обеспечивающих
относительное динамическое постоянство
внутренней среды и устойчивость основных
физиологических функций.
Функциональный антагонизм симпатического и парасимпатического отделов вегетативной нервной системы. Большинство внутренних органов имеет двойную симпатическую и парасимпатическую иннервацию. Антагонизм: сердце. Синергизм: слюнные железы.
+ Медиаторы ВНС. Рецепторы ВНС. Классификация вегетативных рефлексов. Зоны Захарьина-Геда. Механизм действия катехоламинов и ацетилхолина.
Ацетилхолин высвобождается в окончаниях холинергических парасимпатических и симпатических волокон. Инактивация медиатора происходит с помощью фермента ацетилхолинэстеразы.
Н-холинорецепторы в периферических отделах вегетативной нервной системы расположены в ганглионарных синапсах симпатического и парасимпатического отделов, в каротидных клубочках и хромаффинных клетках мозгового слоя надпочечников. Возбуждение этих холинорецепторов сопровождается соответственно облегчением проведения возбуждения через ганглии, что ведет к повышению симпатического и парасимпатического тонуса; повышением рефлекторного возбуждения дыхательного центра, в результате чего углубляется дыхание. Н-холино-рецепторы подразделили на Н-холинорецепторы ганглионарного типа (Нн-холинорецепторы) и мышечного типа (Нм-холинорецепторы). Никотиновые холинорецепторы относятся к ионотропным (быстрым) рецепторам, т.е. они сами выполняют и функцию рецепторов и одновременно являются ионными каналами для катионов (Na+ и К+). Н-холинорецепторы предназначены опосредовать быстрые и непродолжительные эффекты.
М-холинорецепторы блокируются атропином. Все М-холинорецепторы относятся к метаботропному типу. М-холинорецепторы реагируют медленно и длительно.
 М1-холинорецепторы кроме ЦНС обнаружены
также на хромаффиноподобных клетках
желудочных желез.
М1-холинорецепторы кроме ЦНС обнаружены
также на хромаффиноподобных клетках
желудочных желез.
М2-холинорецепторы располагаются в проводящей системе сердца.
М3-холинорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов и экзокринных железах.
Внесинаптические М3-холинорецепторы, которые располагаются в эндотелии сосудов, где они ассоциированы с сосудорасширяющим фактором – окисью азота. Их возбуждение приводит к расширению сосудов и понижению артериального давления.
Н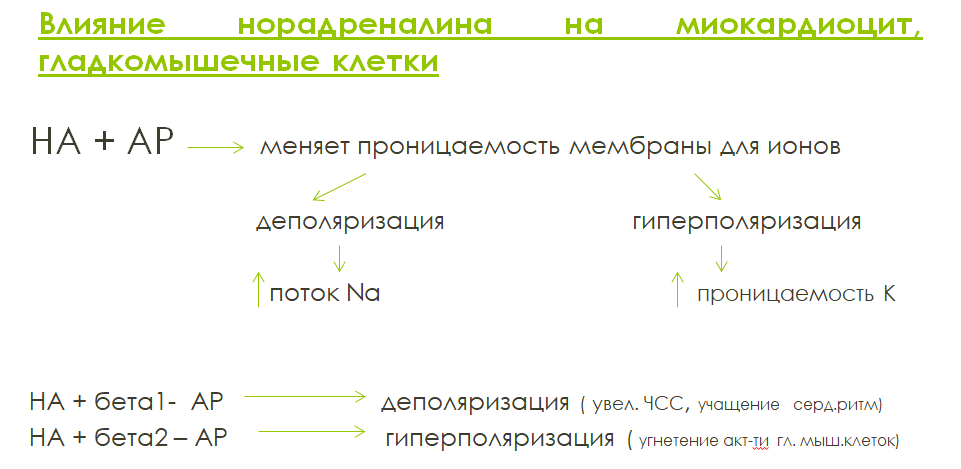 орадреналин
обеспечивает химическую передачу
нервного импульса в адренергических
синапсах вегетативной нервной системы.
Норадреналин относится к катехоламинам.
Главный механизм инактивации норадреналина
– это обратный захват пресинаптической
мембраной с последующим повторным
использованием. Небольшая часть
разрушается ферментами
катехол-О-метилтрансферазой (КОМТ)
в области постсинаптической мембраны
и моноаминоксидазой (МАО), локализованной
на пресинаптической мембране.
Адренорецепторы делят на α-адренорецепторы
и β-адренорецепторы. α- и β1-адренорецепторы
обладают одинаковой чувствительностью
как к норадреналину, так и к адреналину.
β2- и β3- адренорецепторы более чувствительны
к адреналину. Все адренорецепторы
относятся к метаботропному пути.
орадреналин
обеспечивает химическую передачу
нервного импульса в адренергических
синапсах вегетативной нервной системы.
Норадреналин относится к катехоламинам.
Главный механизм инактивации норадреналина
– это обратный захват пресинаптической
мембраной с последующим повторным
использованием. Небольшая часть
разрушается ферментами
катехол-О-метилтрансферазой (КОМТ)
в области постсинаптической мембраны
и моноаминоксидазой (МАО), локализованной
на пресинаптической мембране.
Адренорецепторы делят на α-адренорецепторы
и β-адренорецепторы. α- и β1-адренорецепторы
обладают одинаковой чувствительностью
как к норадреналину, так и к адреналину.
β2- и β3- адренорецепторы более чувствительны
к адреналину. Все адренорецепторы
относятся к метаботропному пути.
Д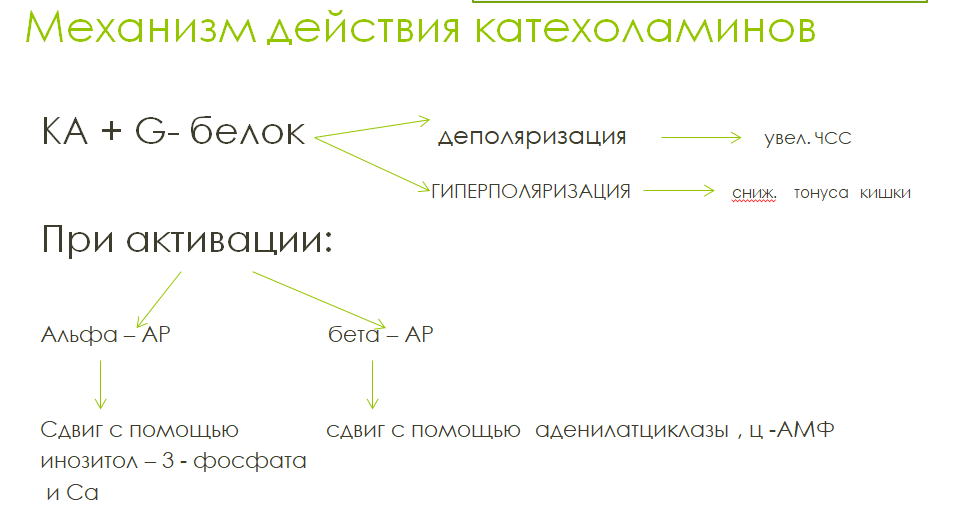 офамин
осуществляет химическую передачу
нервных импульсов не только в
дофаминергических синапсах ЦНС, но и
во вставочных нейронах симпатических
ганглиев и во внутриорганном отделе
вегетативной нервной системы. Инактивация
дофамина осуществляется ферментами
КОМТ и МАО, а также путем обратного
нейронального захвата. В целом на органы
схожее действие с ПНС.
офамин
осуществляет химическую передачу
нервных импульсов не только в
дофаминергических синапсах ЦНС, но и
во вставочных нейронах симпатических
ганглиев и во внутриорганном отделе
вегетативной нервной системы. Инактивация
дофамина осуществляется ферментами
КОМТ и МАО, а также путем обратного
нейронального захвата. В целом на органы
схожее действие с ПНС.
А ТФ
локализация пресинаптические терминали
эффекторных нейронов внутриорганного
отдела вегетативной нервной системы.
Эта передача получила название
пуринергической, так как при стимуляции
этих окончаний выделяются пуриновые
продукты распада – аденозин и инозин.
Действие АТФ проявляется в основном в
расслаблении гладкой мускулатуры.
Пуринергические нейроны являются,
по-видимому, главной антагонистической
тормозной системой по отношению к
холинергической возбуждающей системе.
ТФ
локализация пресинаптические терминали
эффекторных нейронов внутриорганного
отдела вегетативной нервной системы.
Эта передача получила название
пуринергической, так как при стимуляции
этих окончаний выделяются пуриновые
продукты распада – аденозин и инозин.
Действие АТФ проявляется в основном в
расслаблении гладкой мускулатуры.
Пуринергические нейроны являются,
по-видимому, главной антагонистической
тормозной системой по отношению к
холинергической возбуждающей системе.
Серотонин оказывает свое влияние путем взаимодействия со специфическими серотониновыми рецепторами. Периферические S1-рецепторы (или 5-НТ1) в основном обнаружены в гладких мышцах желудочно-кишечного тракта, сосудах скелетных мышц и сердца, проводящей системе сердца. Действие схоже с СНС.
А спарагиновая
и глутаминовая кислоты являются
медиаторами возбуждающего типа,
гамма-аминомасляная кислота –
медиатором тормозного типа. В результате
взаимодействия ГАМК с ГАМК-рецепторами
происходит открытие каналов для ионов
хлора, что обусловливает гиперполяризующее
действие ГАМК.
спарагиновая
и глутаминовая кислоты являются
медиаторами возбуждающего типа,
гамма-аминомасляная кислота –
медиатором тормозного типа. В результате
взаимодействия ГАМК с ГАМК-рецепторами
происходит открытие каналов для ионов
хлора, что обусловливает гиперполяризующее
действие ГАМК.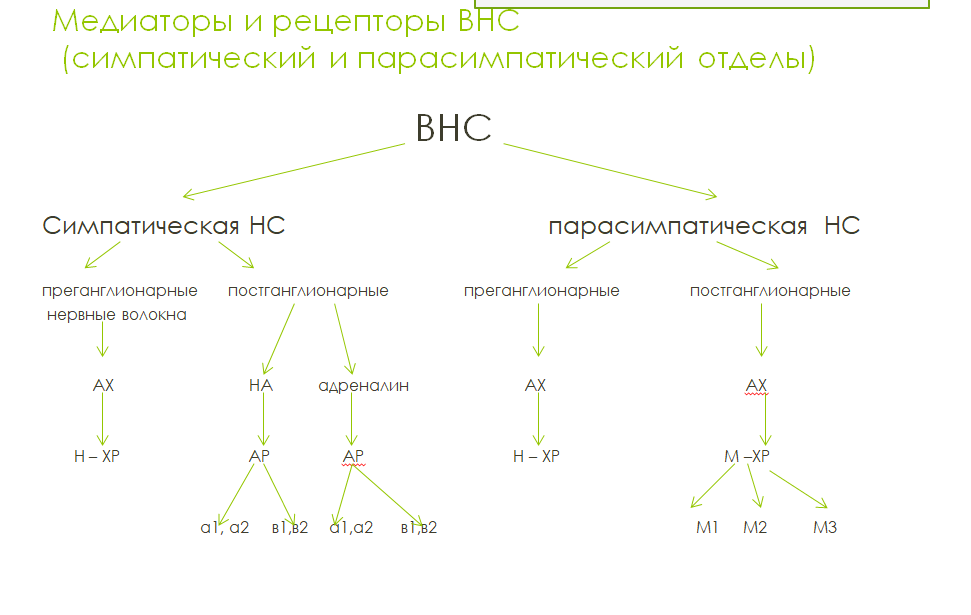
Классификация вегетативных рефлексов:
Висцеровисцеральные. Классическим примером висцеро-висцерального рефлекса является рефлекс Гольца, показывающий, что механическое раздражение брыжейки вызывает замедление частоты сердечных сокращений. Разновидностью висцеро-висцерального рефлекса является аксон-рефлекс, например, возникновение сосудистой реакции при раздражении кожных болевых рецепторов.
К висцеросоматическим рефлексам относятся торможение общей двигательной активности организма при раздражении хемо- и механорецепторов синокаротидной зоны, а также сокращение мышц брюшного пресса или подергивание конечностей при раздражении рецепторов пищеварительного тракта.
При висцеросенсорных рефлексах в ответ на раздражение вегетативных чувствительных волокон возникают не только реакции во внутренних органах, но и изменяется соматическая чувствительность. Для его вызова необходимо продолжительное и сильное воздействие. Зона повышенного восприятия обычно ограничивается участком кожи, иннервируемым сегментом, к которому поступают импульсы от раздражаемого висцерального органа. В клинике имеют определенное значение висцеродермальные рефлексы. Вследствие сегментарной организации вегетативной и соматической иннервации при заболеваниях внутренних органов на ограниченных участках кожи возникает повышение тактильной и болевой чувствительности. Эти боли называют отраженными, а области, в которых они появляются, – зонами Захарьина–Геда.
При раздражении некоторых областей поверхности тела возникают сосудистые реакции и изменения функций определенных висцеральных органов. Это дермовисцеральный рефлекс, который является разновидностью соматовисцерального рефлекса. На этом основана рефлексотерапия.
+ Спинальные центры регуляции вегетативных функций. Гипоталамические центры регуляции. Мозжечковые центры и центры ретикулярной формации. Центры лимбической системы и коры большого мозга. Факторы, обеспечивающие тонус вегетативных центров.
Спинальные центры регуляции вегетативных функций. В грудной части спинного мозга (в боковых рогах) находятся тела нейронов симпатической НС. В крестцовом отделе находятся центры эвакуаторных рефлексов, относящиеся к парасимпатической НС.
Гипоталамические центры регуляции. Гипоталамус регулирует гомеостаз, интегративные функции, такие как температуре тела, аппетит, потребление воды, мочеиспускание, сердечную деятельность, сексуальную активность и рост человека.
Ключевая роль гипоталамуса в регуляции вегетативных функций объясняется несколькими факторами. Во-первых, гипоталамус получает обширную информацию от разных структур: от интерорецепторов, которая идёт через ствол мозга, от экстерорецепторов (слуховых, поверхности кожи). Некоторые нейроны гипоталамуса сами имеют рецепторы, которые могут оценивать состав крови, ЦСЖ, давление и температуру. В гипоталамусе находятся терморецепторы, глюкорецепторы, осморецепторы. Во-вторых, гипоталамус имеет многочисленные связи с лимбической системой, с РФ, с корой больших полушарий. Эти связи лежат в основе координации вегетативных функций с определённым поведением, например, с эмоциями. В-третьих, гипоталамус образует проекции на вегетативные центры ствола и спинного мозга, т.е. осуществляет прямой контроль над нижележащими вегетативными центрами. В-четвёртых, гипоталамус регулирует эндокринную систему организма.
Гипоталамус регулирует околосуточные (циркарные) ритмы. Многие вегетативные функции находятся под влиянием биологических часов: концентрация гормонов, перепады температуры тела, повышение и снижение работоспособности. Ключевая структура для генерации суточного ритма в гипоталамусе – супрахиазматическое ядро. К этому ядру подходят волокна от зрительных нервов. Если это ядро разрушить, теряется возможность настраивать физиологические ритмы в соответствии с ритмами дня и ночи.
При раздражении задней части гипоталамуса у животных активируется симпатическая часть НС. Раздражение передней части гипоталамуса активирует парасимпатическую часть НС.
Нейроны гипоталамуса помимо участия в процессах регуляции водно-солевого поведения, лактации, активности матки, способны продуцировать гормоны (окситоцин и ЛДГ). Помимо секреции этих гормонов, способны управлять выделением этих гормонов. Часть нейронов гипоталамуса продуцирует пептидные гормоны (либерины и статины).
Мозжечковые центры регуляции. Принимает участие в регуляции функций внутренних органов, стабилизирует гомеостазис – при его удалении он становится неустойчивым, т.е. угнетается активность кишечных желез и моторики ЖКТ. Раздражение мозжечка вызывает сужение кровеносных сосудов, расширение зрачка, учащение сердцебиений, изменение дыханий и т.д.
РФ центры регуляции – управляет вегетативными функциями посредством активации симпатической НС, ее нейроны формируют центры продолговатого мозга (дыхательный, кровообращения).
Лимбическая система. Благодаря гипоталамусу все эмоциональные реакции, которые реализуются с участием лимбической системы, приобретают конкретную эндокринную окраску.
КБП. Высший интегративный центр регуляции всех вегетативных функций организма. Стимуляция двигательной доли коры вызывает изменение деятельности ССС (увеличение минутного объема сердца, усиление кровообращения в мышцах). Раздражение лобных долей ведет к изменению сердечной деятельности и дыхательного ритма, но и даже может изменить деятельность любого органа, имеющего вегетативную иннервацию.
Факторы, обеспечивающие тонус вегетативных центров.
Спонтанная активность нейронов того или иного центра (РФ);
Поток афферентных импульсов в ЦНС от различных рефлексогенных зон (проприорецепторы);
Действия БАВ и метаболитов на клетки центра (СО2)
Значение тонуса вегетативных центров заключается в том, что один и тот же центр с помощью одних и тех же эфферентных нервных волокон может вызвать двоякий эффект деятельности органов.
+ Функции спинного мозга. Нейроны спинного мозга. Миостатические и позно-тонические рефлексы. Синдром Броун-Секара. Спинальный шок.
Функции спинного мозга.
Рефлекторная: осуществляется нервными центрами спинного мозга, которые являются сегментарными рабочими центрами безусловных рефлексов. Их нейроны сообщаются с рецепторами и рабочими органами. Каждый метамер (поперечный участок) тела получает чувствительность от 3 корешков. Скелетные мышцы также получают иннервацию от 3 соседних сегментов спинного мозга. Эфферентные импульсы идут к скелетной мускулатуре, дыхательным мышцам, внутренним органам, сосудам и железам. Вышележащие отделы ЦНС управляют переферией с помощью сегментарных отделов спинного мозга.
Проводниковая: осуществляется за счет восходящих и нисходящих путей спинного мозга. Восходящие пути передают информацию от тактильных, болевых, температурных и проприорецепторов мышц и сухожилий через нейроны спинного мозга в другие отделы ЦНС к мозжечку и коре большого мозга.
Нейроны спинного мозга.
В первую группу входят мотонейроны, или двигательные нейроны, расположенные в передних рогах, а их аксоны образуют передние корешки спинномозговых нервов. Альфа-мотонейроны иннервируют экстрафузальные мышечные волокна, обеспечивая мышечное сокращение. На дендритах этих клеток располагается около 20 тысяч синапсов. Альфа-мотонейроны обладают низкой частотой импульсации (10–20 имп/с). Гамма-мотонейроны иннервируют интрафузальные мышечные волокна мышечного веретена. Они обладают высокой частотой импульсации (до 200 имп/с). Главная функция гамма-мотонейронов состоит в предотвращении во время сокращения экстрафузальных волокон перерастяжения мышцы. Возбуждение гамма-мотонейронов способствует развитию начавшегося движения и используется для выполнения особенно тонких и мелких движений.
Вторую группу составляют интернейроны – промежуточные нейроны, расположенные в задних рогах и получающие информацию от чувствительных спинномозговых ганглиев. Они реагируют на болевые, температурные, тактильные и проприоцептивные раздражители. Интернейроны отвечают за висцеромоторные рефлексы. Кроме того, они обеспечивают восходящие и нисходящие тормозные и возбуждающие влияния на клетки выше- и нижележащих структур спинного мозга. Интернейроны участвуют в реципрокном торможении.
Третья группа нейронов – это симпатические и парасимпатические нейроны, расположенные в боковых рогах. Аксоны этих клеток выходят из спинного мозга в составе передних корешков спинномозговых нервов. Разряды симпатических нейронов синхронизируются с колебаниями артериального давления.
К четвертой группе относят ассоциативные нейроны, обеспечивающие внутри- и межсегментарные связи. В средней части спинного мозга находится промежуточное ядро, содержащее нейроны, аксоны которых образуют сеть – студенистое вещество (желатинозную субстанцию Роланда), или ретикулярную формацию спинного мозга.
Альфа- и гамма-мотонейроны поддерживают тонус тела и обеспечивают рефлексы сгибания и разгибания – миостатические рефлексы, или рефлексы на растяжение мышцы. Механический удар неврологическим молоточком по сухожилию мышцы приводит к быстрому ее растяжению всего на несколько миллиметров, а затем сокращению всей мышцы и появлению ответной реакции. К таким рефлексам относятся: коленный, ахиллов, подошвенный, сгибательный и разгибательный рефлексы предплечья.
Познотонические рефлексы спинного мозга направлены на поддержание позы. С их помощью регулируется тонус мышц. Эти рефлексы возникают с проприорецепторов мышц шеи и регулируют тонус мышц при изменении положения головы и шеи.
При одностороннем повреждении см (синдром Броун–Секара). На стороне поражения (ниже места поражения) нарушаются двигательные функции вследствие повреждения пирамидного тракта, а также проприоцептивная чувствительность. На противоположной стороне движения сохраняются, но отсутствует болевая и температурная чувствительность (повреждение перекрещенного спиноталамического тракта) и с обеих сторон частично нарушена тактильная чувствительность.
Полная перерезка или повреждение спинного мозга приводит к возникновению спинального шока (шок-удар). В результате ниже перерезки исчезают все виды рефлекторной деятельности: нарушается двигательная активность, все виды чувствительности, вегетативные функции. Мочеиспускание и отделение кала становятся непроизвольными, наступает расширение сосудов, падение артериального давления, нарушается процесс теплообразования, увеличивается теплоотдача.
+ Функции продолговатого мозга и варолиева моста. Функции среднего мозга. Децеребрационная ригидность. Нистагм головы и глаз. Рефлекс «лифта».
Функции продолговатого мозга:
Собственные. В продолговатом мозге расположены ядра VIII (слух), IX (чувствительность, движения глотки и языка), X, XI (ГКС и трапециевидная) и XII (язык – сосание, глотание, жевание, речь) пар черепно-мозговых нервов.
Вегетативные центры: дыхание, ССС, сердечной деятельности, слюноотделения. Продолговатый мозг отвечает за некоторые защитные рефлексы: рвоты, кашля, чихания, слезоотделения, смыкания век, а также рефлексы пищевого поведения (сосания, жевания, глотания). Продолговатый мозг реализует соматические рефлексы: рефлексы поддержания позы тела за счет статических рефлексов, регулирующих тонус мышц, удерживающих положение тела в пространстве в покое.
Проводниковые. В продолговатом мозге начинаются нисходящие пути: вестибулоспинальный, оливоспинальный, ретикулоспинальный, обеспечивающие связь между вестибулярными ядрами, оливой, ретикулярной формацией продолговатого мозга и мотонейронами спинного мозга, отвечающими за тонус и координацию мышечных реакций. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спиноталамический и спинномозжечковый путь; пирамидальный, руброспинальный. В нем заканчиваются пути восходящие пути проприоцептивной чувствительности – тонкий пучок Голля и клиновидный пучок Бурдаха.
Поэтому при повреждении одной из половин продолговатого мозга выше перекреста восходящих путей проприоцептивной чувствительности происходит нарушение чувствительности и работы мышц лица и головы на стороне повреждения, в то время как на противоположной стороне возникают нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это связано с тем, что все восходящие и нисходящие пути перекрещиваются.
Функции моста:
Собственные функции: ядро 8 (вестибулярный), ядро 7 (лицевая мускулатура, стременная мышца, вкус), 6 (отводит глаз), 5 (жевательная, тактильная и тд; участвует в чихательном, жевательном и глотательном рефлексах.).
В мосте располагается пневмотаксический центр. Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга. Она влияет на кору больших полушарий, активируя ее и вызывая реакцию пробуждения. Аксоны ретикулярной формации моста идут в мозжечок и спинной мозг.
Проводниковая функция мозга. Через мост проходят все восходящие и нисходящие пути, связывающие мост с мозжечком и спинным мозгом, корой больших полушарий и другими структурами центральной нервной системы.
Функции среднего мозга:
Собственные функции: ядро блокового нерва – IV пара черепно-мозговых нервов (движение глаз), ядро глазодвигательного нерва (движение глаз + слезная железа), красное ядро (тонус мускулатуры), черное вещество (регуляции актов жевания, глотания и их последовательности, а также в координации мелких и точных движений пальцев рук), четверохолмие, ножки мозга и ядра ретикулярной формации (сон и бодростовование).
Во время фазы «быстрого сна» повышается активность нейронов голубого пятна, которые с помощью норадреналина выключают мышечный тонус и фазические движения мускулатуры туловища и конечностей.
Проводниковая функция среднего мозга определяется наличием в нем как восходящих путей к таламусу (спиноталамический тракт, медиальная петля), к коре больших полушарий и мозжечку, так и нисходящих – к продолговатому и спинному мозгу (пирамидный тракт, корково-мостовой).
Рефлекторная. Верхние бугры четверохолмия – это первичный зрительный подкорковый центр, обеспечивающий зрительный ориентировочный рефлекс – поворот головы и глаз в сторону светового раздражителя, фиксацию взора и слежение за движущимися объектами. При повреждении или раздражении верхних бугров наблюдается нистагм глаз – непроизвольные ритмические и колебательные движения глаз (горизонтальный, вертикальный, вращательный).
Нижние бугры четверохолмия – это первичный слуховой подкорковый центр, участвующий в ориентировочном слуховом рефлексе – повороте головы в сторону источника звука. Двустороннее повреждение нижних бугров сопровождается потерей способности локализовать источник звука в пространстве.
Децеребрационная ригидность — повышенние тонуса мышц разгибателей и относительное расслабление мышц-сгибателей, возникает в результате перерезания стволовой части головного мозга (на уровне среднего мозга) — децеребрации, объясняется прекращением тормозящего действия красного ядра на ядро Дейтерса. В эксперименте на кошках, при перерезании у них структур, соединяющих вышеуказанные ядра, животные не могут выполнить сгибание.
Средний мозг осуществляет и стато-кинетические рефлексы. Это рефлексы, которые служат для сохранения устойчивого положения тела при движении. Лифтная реакция – это уменьшение тонуса разгибателей конечностей в начале быстрого подъема, которое сменяется его повышением. При быстром опускании, тонус разгибателей меняется противоположным образом.
+ Понятие «висцерального мозга». Интегративная деятельность гипоталамуса. Роль лимбической системы. Интегративная деятельность мозжечка. Интегративная деятельность продолговатого мозга.
Интегративная деятельность продолговатого мозга. В его нейронах запрограммированы алгоритмы некоторых сложных регуляторных процессов, требующие для своего осуществления участия центров других отделов нервной системы и взаимодействия с ними. Например, компенсационное изменение положения глаз при колебаниях головы во время движения, реализуемое на основе взаимодействия ядер вестибулярной и глазодвигательной систем мозга с участием медиального продольного пучка. Часть нейронов ретикулярной формации продолговатого мозга обладает автоматией, влияет на дыхательный, сердечный центр и тд.
Интегративная деятельность мозжечка. Мозжечок регулирует позу и равновесие, силу и точность мышечных сокращений, и их тонус, как в покое, так и при движениях, а также синергию сокращений разных мышц при сложных движениях. Координация позы и выполняемых целенаправленных движений осуществляется старым и новым мозжечком (околочревной зоной). Эта область мозжечка получает информацию, как от проприорецепторов двигательного аппарата, так и от моторной коры (программа произвольного движения) и после ее анализа, осуществляемого через промежуточные ядра и красное ядро, проводится координация позы и соответствующая коррекция выполняемого целенаправленного движения. Мозжечок имеет связи с проприорецепторами, кожными и вестибулярными рецепторами, оказывая влияние на сенсорные системы. Он регулирует возбудимость сенсомоторной коры больших полушарий, контролируя тем самым уровень тактильной, температурной и зрительной чувствительности. Предполагают участие мозжечка в процессах мышления (например, при арифметическом счете).
Мозжечок оказывает как угнетающее, так и стимулирующее влияние на работу сердечно-сосудистой системы. При раздражении последнего высокое артериальное давление снижается, а исходное низкое – повышается, увеличивается ЧСС, уменьшается частота дыхания, изменяется тонус гладких мышц кишечника, происходит расширение зрачка. Стимуляция мозжечка приводит к нарушению репродуктивной функции, к сокращению матки у беременных кошек. Удаление мозжечка провоцирует рождение нежизнеспособного потомства или препятствует зачатию. Предполагают, что за вегетативные функции отвечает старый и древний мозжечок (преимущественно червь), получающий информацию от интерорецепторов различных органов.
Интегративная деятельность гипоталамуса. Гипоталамус является структурой ЦНС, осуществляющей сложную интеграцию и приспособление функций различных внутренних органов к целостной деятельности организма. Гипоталамус объединяет и связывает в единое целое механизмы гуморальной и нервной регуляции. Под контролем гипоталамуса находятся такие железы внутренней секреции, как гипофиз, щитовидная, половые железы, надпочечники и др. Регуляция тропных функций гипофиза осуществляется путем выделения гипоталамическими нейронами гормонов, поступающих в гипофиз в основном через портальную систему сосудов. Выделение тропных гормонов гипофиза приводит к изменению функций эндокринных желез, секрет которых попадает в кровь и в свою очередь может действовать на гипоталамус (обратная связь). Передняя область гипоталамуса оказывает стимулирующее влияние на половое развитие организма. Гормоны нейрогипофиза являются продуктом секреции супраоптического ядра гипоталамуса (например, вазопрессин или антидиуретический гормон). Под контролем гипоталамических центров находятся такие интегративные функции организма, как поддержание постоянства температуры тела, углеводный, жировой и водный обмены организма, регуляция давления крови, регуляция половых функций и функций желудочно-кишечного тракта и др.
Лимбическая система. Лимбическая система – это функционально единый комплекс нервных структур, ответственных за эмоциональное поведение, побуждения к действию (мотивации), процессы научения и запоминания, инстинкты (пищевые, оборонительные, половые) и регуляцию цикла «сон–бодрствование». В связи с тем, что лимбическая система воспринимает большое количество информации от внутренних органов, она получила также название «висцеральный мозг».
В так называемый круг Пейпеса входят: гиппокамп – мамиллярные тела – передние ядра таламуса – кора поясной извилины – парагиппокампальная извилина – гиппокамп. Этот круг отвечает за эмоции, формирование памяти и обучение.. Лимбическая система помимо связей с ассоциативной корой образует связи с лобной и височной долями. Последние передают информацию от зрительной, слуховой и соматосенсорной коры к миндалине и гиппокамп.
+ Физиология базальных ганглий. Физиология ретикулярной формации. Структурно-функциональные особенности коры большого мозга. Локализация функций в коре больших полушарий. Электрическая активность коры головного мозга.
Физиология базальных ганглий. Базальные ганглии, или подкорковые ядра, относятся к структурам переднего мозга и включают в себя полосатое тело (corpus striatum), или неостриатум: хвостатое ядро (n.caudatus) и скорлупу (putamen), палеостриатум (бледный шар, или globus pallidus) и ограду (claustrum). Их поражение вызывает болезнь Паркинсона.
Помимо афферентных и эфферентных связей базальных ганглиев, в их структуре выделяют несколько параллельных функциональных петель, связывающих кору больших полушарий (двигательную, соматосенсорную и лобную) с таламусом.
Информация поступает из фронтальных зон коры, проходит через базальные ядра (хвостатое ядро и скорлупу) и черное вещество в двигательные ядра таламуса, оттуда снова возвращается в эти же зоны коры – это скелетомоторная петля (управляет движениями лица и рта).
Другая петля – глазодвигательная (окуломоторная) соединяет поле 8 лобной коры с хвостатым ядром. Далее импульсация поступает в бледный шар и черное вещество, проецируясь в вентральное переднее (VA) ядро таламуса, а затем возвращается снова в поле 8 лобной коры.
Скорлупа отвечает за организацию пищевого поведения. При ее поражении наблюдаются трофические нарушения кожи, а ее раздражение вызывает слюноотделение и изменение дыхания. Функции бледного шара заключаются в провоцировании ориентировочной реакции, движения конечностей, пищевого поведения (жевание, глотание).
Физиология ретикулярной формации. Реетикулярная формация (РФ) располагается в центральной части мозгового ствола, заходя ростральным концом в таламус, а каудальным – в спинной мозг. Благодаря наличию сетевых связей почти со всеми структурами центральной нервной системы она получила название ретикулярной, или сетевой, формации.
Ретикулярная формация оказывает генерализованное, тонизирующее, активирующее влияние на передние отделы головного мозга и кору больших полушарий (восходящая активирующая система РФ), и нисходящее, контролирующее деятельность спинного мозга (нисходящая ретикулоспинальная система), которое может быть как облегчающим на многие функции организма, так и тормозным. Одним из видов тормозного влияния РФ на рефлекторную деятельность спинного мозга является «сеченовское торможение», заключающееся в угнетении спинальных рефлексов при раздражении таламической ретикулярной формации кристалликом соли. Раздражение ретикулярной формации вызывает тремор, спастический тон.
РФ среднего мозга играет роль в координации сокращений глазных мышц. Получив информацию от верхних бугров четверохолмия, мозжечка, вестибулярных ядер, зрительных областей коры головного мозга, РФ ее интегрирует, что приводит к рефлекторным изменениям работы глазодвигательного аппарата, особенно при внезапном появлении движущихся объектов, изменении положения головы и глаз.
В ретикулярной формации продолговатого, среднего мозга и таламуса имеются нейроны, реагирующие на болевые раздражения от мышц и внутренних органов, при этом создается ощущение тупой боли.
Физиология коры больших полушарий.
Кора большого мозга представляет собой многослойную нейронную ткань с множеством складок. В коре выделяют 6 слоев (I—VI), каждый из которых состоит из пирамидных и звездчатых клеток. Кортикализация функций — возрастание в филогенезе роли коры большого мозга в анализе и регуляция функций организма и подчинение себе нижележащих отделов ЦНС. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов, которые у других млекопитающих сохраняются.
Локализация функций в коре больших полушарий.
Принято делить на сенсорные, двигательные и ассоциативные.
В сенсорных зонах представлены корковые концы всех анализаторов.
Для зрительного он располагается в затылочной доле мозга (поля 17, 18, 19). В поле 17 заканчивается центральный зрительный путь, информирующий о наличии и интенсивности зрительного сигнала. Это первичная зрительная область, в которой имеется топическое представительство рецепторов сетчатки, при этом каждому участку сетчатки соответствует свой участок зрительной коры. Поля 18 и 19 – это вторичная зрительная область, которая анализирует цвет, форму, размеры и качество предмета.
Корковый конец слухового анализатора локализуется в височной доле коры (извилина Гешля), полях 41, 42, 22. Они участвуют в восприятии и анализе слуховых раздражений по их громкости и тональности, организации слухового контроля речи, за который отвечает слуховой центр речи Вернике, расположенный в задних отделах верхней височной извилины (поля 22). Этот центр отвечает за распознавание и хранение устной речи как собственной, так и чужой (речевой гнозис).
В височной доле располагается также корковый конец вестибулярного анализатора (задний отдел поля 21).
Кожный анализатор, а также болевая, температурная и проприоцептивная чувствительность проецируются на заднюю центральную извилину (поля 1, 2, 3, 43), в верхней части которой представлены нижние конечности, в средней – туловище, в нижней – руки и голова.
Задняя центральная извилина вместе с прецентральной, в которой располагаются моторные зоны, объединены в так называемую «соматосенсорную кору», поскольку кожный и двигательный анализаторы тесно связаны анатомически и образуют единую функциональную единицу, обеспечивающую двигательную активность живого организма.
Корковый конец обонятельного и вкусового анализаторов расположен в гиппокампальной извилине (поля 34, 43).
Моторные зоны находятся в лобных долях. Кора прецентральной извилины (поле 4) представляет первичную двигательную зону. В пятом слое этого поля находятся очень крупные пирамидные клетки (гигантские клетки Беца), формирующие эфферентные выходы моторной коры на стволовые и спинальные центры. Премоторная область коры (поле 6) располагается кпереди от первичной двигательной зоны. Поле 6 называют вторичной моторной областью, функции которой связаны с планированием и координацией произвольных движений. В премоторной области коры располагается центр письменной речи (задний отдел средней лобной извилины – поле 6), центр моторной речи Брока (задний отдел нижней лобной извилины левого полушария – поле 41), а также музыкальный моторный центр (поле 45), отвечающий за тональность речи и способность петь. К полю 6 примыкает поле 8 (лобное глазодвигательное), раздражение которого сопровождается поворотом головы и глаз в сторону, противоположную раздражаемой.
К ассоциативной коре относят теменно-височно-затылочную, префронтальную и лимбическую области. Теменная ассоциативная область коры больших полушарий (поля 5, 7, 40) участвует в формировании субъективного представления об окружающем пространстве, о нашем теле («схеме тела»), узнавании (гнозисе) предметов, их распознавании. Височная область коры участвует в речевой функции посредством слухового контроля речи (слуховой центр речи Вернике). Лобные ассоциативные поля (поля 9–14) имеют прямое отношение к лимбическим отделам мозга, они принимают участие в формировании программы целенаправленных, сложных поведенческих актов в ответ на воздействие внешней среды на основе сенсорных сигналов всех модальностей, особенно в новой для человека обстановке.
Электрическая активность коры головного мозга.
Мембранный потенциал пирамидных клеток составляет от 50 до 80 мкВ, потенциал — действия 60—100 мкВ. Частота ПД — около 100 Гц.
Ритмы мозговой активности на ЭЭГ:
Альфа-ритм характеризуется частотой колебаний 8-13 Гц, амплитуда 50 мкВ. Этот ритм лучше всего выражен в затылочной и теменной областях коры и регистрируется в условиях физического и умственного покоя при закрытых глазах. Если глаза открыть, то альфа-ритм сменяется более быстрым бета-ритмом.
Бета-ритм характеризуется частотой колебаний 14-50 Гц и амплитудой до 15 мкВ.
Тета-ритм представляет собой колебания с частотой 4-8 Гц и амплитудой 100-150 мкВ. Этот ритм регистрируется, во время поверхностного сна, при гипоксии и легком наркозе.
Дельта-ритм характеризуется медленными колебаниями потенциалов с частотой 0,5-3,5 Гц, амплитудой 250-300 мкВ. Этот ритм регистрируется во время глубокого сна, при глубоком наркозе, при коматозном состоянии.
+ Гормоны и их свойства. Функции гормонов. Классификация гормонов. Варианты действия гормонов. Регуляция секреции гормонов.
Гормоны – это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами, и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма.
Свойства гормонов:
1) Органоспецифичность;
2) Высокая биологическая активность;
3) Дистантный характер действия;
4) Генерализованность действия;
5) Пролонгированность действия.
6) Строгая специфичность – 1 гормон нельзя заменить другим.
Функции гормонов
1. Регуляция роста, развития и дифференцировки тканей и органов, что определяет физическое, половое и умственное развитие.
2. Обеспечение адаптации организма к меняющимся условиям существования. Например, при адаптации к холоду, изменению уровня физической активности, характеру питания, беременности, родам, вскармливанию и т.д.
3. Обеспечение гомеостаза. Например, регуляция содержания глюкозы в крови, концентрации ионного состава, общего объема циркулирующей жидкости, осмотического давления и т.д.
Классификация гормонов и БАВ по химической структуре
Производные аминокислот:
• производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин;
производные триптофана: мелатонин, серотонин;
производные гистидина: гистамин.
Белково-пептидные гормоны:
• полипептиды: глюкагон, кортикотропин, меланотропин, вазопрессин, окситоцин, пептидные гормоны желудка и кишечника;
• простые белки (протеины): инсулин, соматотропин, пролактин, паратгормон, кальцитонин;
• сложные белки (гликопротеины или гликопротеиды): тиротропин, фоллитропин, лютотропин.
Стероидные гормоны:
• кортикостероиды (альдостерон, кортизол, кортикостерон);
• половые гормоны: андрогены (тестостерон), эстрогены и прогестерон.
Производные жирных кислот:
• арахидоновая кислота и ее производные: простагландины, простациклины, тромбоксаны, лейкотриены.
Функциональная классификация гормонов
1. Рилизинг-гормоны – гормоны, регулирующие синтез и выделение гормонов аденогипофиза, преимущественно тропных. Синтезируются и выделяются нейросекреторными клетками гипоталамуса.
2. Тропные гормоны – гормоны, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Образуются и выделяются аденогипофизом.
3. Эффекторные гормоны – гормоны, которые оказывают влияние непосредственно на орган-мишень. Образуются и выделяются в основном периферическими железами внутренней секреции, а также гипофизом.
Варианты действия гормонов:
Гормональное (собственно эндокринное) – гормон выделяется из клетки – продуцента, поступает в кровь и с током крови подходит к органу – мишени, действуя на расстоянии от места продукции гормона;
Паракринное – из места синтеза гормон попадает во внеклеточное пространство, а из него воздействует на клетки – мишени;
Изокринное – как паракринное, контакт клетки – продуцента гормона и клетки – мишени очень тесный;
Нейрокринное – как и действие медиатора;
Аутокринное - клетка продуцирует гормон, который сам и воздействует на эту же клетку – продуцент.
Регуляция секреции гормонов:
Гормональная – в гипоталамусе вырабатываются либерины и статины, которые через портальную систему гипофиза из гипоталамуса попадают в аденогипофиз и усиливают (либерины) или тормозят (статины) продукцию соответствующих гормонов (АКТГ, ЛГ, СТГ, ТТГ).
Регуляция продукции гормонов по типу обратной отрицательной связи – продукция тиреоидных гормонов щитовидной железы регулируется тиреолиберином гипоталамуса, воздействующего на аденогипофиз, продуцирующий ТТГ, который повышает продукцию тиреоидных гормонов. Войдя в кровь, Т3 и Т4 воздействуют на гипоталамус и аденогипофиз и тормозят продукцию ТТГ и тиреолиберина.
Регуляция с участием структур ЦНС - активность супрахиазматического ядра гипоталамуса вместе с активностью гипофиза обеспечивают существование биологических часов для гормональной секреции.
+ Виды взаимодействия гормонов. Механизм действия гормонов. Синтез белковых гормонов. Синтез стероидных гормонов. Синтез катехоламинов.
Виды взаимодействия гормонов
Синергизм, т.е. однонаправленное действие двух или нескольких гормонов. Например, адреналин и глюкагон активируют распад гликогена печени до глюкозы и вызывают увеличение уровня сахара в крови.
Антагонизм всегда относителен. Например, инсулин и адреналин оказывают противоположное действие на уровень глюкозы в крови. Инсулин вызывает гипогликемию, адреналин – гипергликемию. Биологическое же значение этих эффектов сводится к одному – улучшению углеводного питания тканей.
Пермиссивное действие гормонов заключается в том, что гормон, сам не вызывая физиологического эффекта, создает условия для ответной реакции клетки или органа на действие другого гормона. Например, глюкокортикоиды, не влияя на тонус мускулатуры сосудов и на распад гликогена печени, создают условия, при которых даже небольшие концентрации адреналина увеличивают артериальное давление и вызывают гипергликемию в результате гликогенолиза в печени.
М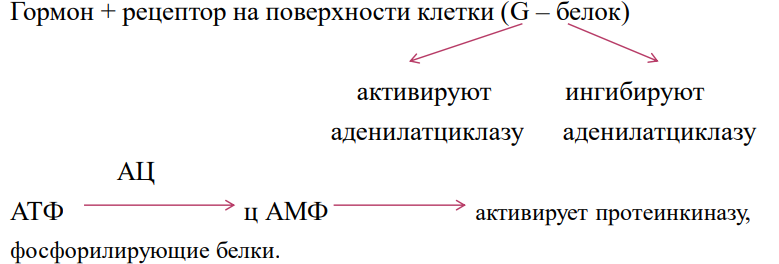 еханизм
действия гормонов
еханизм
действия гормонов
Реализация эффекта с наружной поверхности клеточной мембраны
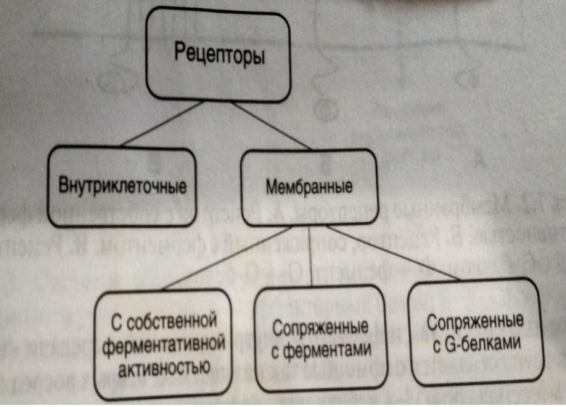
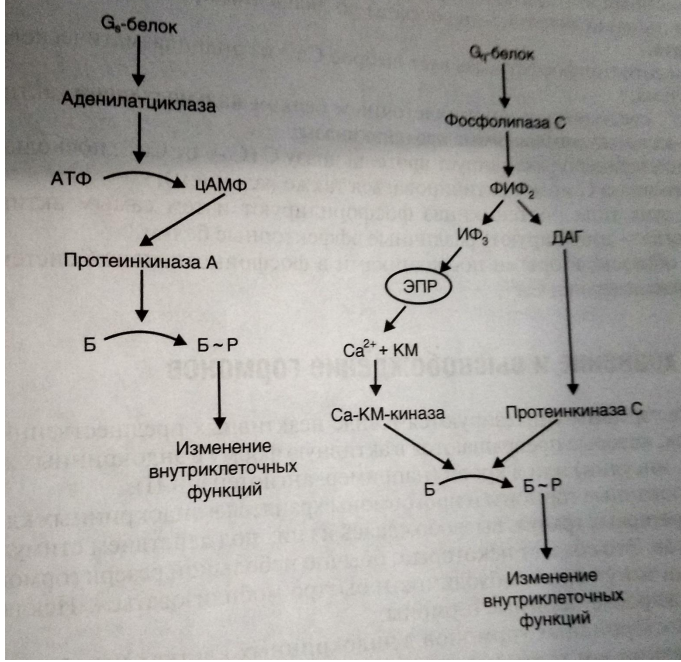
Реализация эффекта после проникновения гормонов внутрь клетки (стероидные гормоны – связывание с рецепторами в цитоплазме, тиреоидные – связывание в ядре). Происходит связывание гормон-рецепторного комплекса с ДНК и белками хроматина, что стимулирует транскрипцию определенных генов. Трансляция мРНК приводит к появлению в клетке новых белков, вызывающих биологический эффект этих гормонов.
Синтез белковых гормонов.
Синтез стероидных гормонов.

Синтез катехоламинов.

+ Гормоны аденогипофиза и их физиологическое значение. Гормоны нейрогипофиза. Тропные гормоны. Эффекторные гормоны. Рилизинг-гормоны.
Гормоны аденогипофиза и их физиологическое значение.
СТГ – принимает участие в регуляции роста, усиливает образование белка. Наиболее выражено его влияние на рост эпифизарных хрящей конечностей, рост костей идет в длину. Влияет на углеводный обмен: стимулирует синтез гликогена и глюкозы, повышает липолиз. Нарушение соматотропной функции гипофиза: если имеется гиперфункция в детском возрасте, то развивается гигантизм; при гипофункции – карликовость. Гиперфункция у взрослого человека не влияет на рост в целом, но увеличиваются размеры тех частей тела, которые еще способны расти (акромегалия). Действие соматотропина происходит посредством соматомединов А и С, которые под влиянием соматотропина образуются в основном в печени. После установления химической структуры соматомединов оказалось, что она напоминает структуру молекулы проинсулина. Поэтому их также назвали инсулиноподобными факторами роста (ИФР-1 и ИФР-2). Секреция соматомединов происходит равномерно на протяжении суток. ИФР-1 (соматомедин С), по-видимому, является основным фактором роста.
Пролактин способствует образованию молока в альвеолах и росту молочных желез, но после предварительного воздействия на них женских половых гормонов. После родов увеличивается синтез пролактина и наступает лактация. Акт сосания через нервно-рефлекторный механизм стимулирует выброс пролактина. Пролактин обладает лютеотропным действием, способствует продолжительному функционированию желтого тела и выработке им прогестерона.
ТТГ увеличивает функцию щитовидной железы, обладает способностью стимулировать образование белка тиреоглобулина в клетках фолликулов щитовидной железы и поступление его в полость фолликула. При сниженной выработке тиреотропина происходит атрофия щитовидной железы, при гиперпродукции – разрастание.
АКТГ - стимулятор пучковой зоны коры надпочечников, где образуются глюкокортикоиды. АКТГ увеличивает синтез холестерина и скорость образования прегненолона из холестерина, вызывает распад и тормозит синтез белка, является антагонистом гормона роста. Он тормозит развитие основного вещества соединительной ткани, уменьшает количество тучных клеток, подавляет фермент гиалуронидазу, снижая проницаемость капилляров. Под влиянием кортикотропина уменьшаются размер и масса лимфоидных органов. Этим определяется его противовоспалительное действие. Секреция кортикотропина подвержена суточным колебаниям: в вечерние часы его содержание выше, чем утром. Секреция АКТГ усиливается при стрессе, а также под влиянием факторов, вызывающих стрессогенные состояния: холод, боль, физические нагрузки, эмоции.
фоллитропин (фолликулостимулирующий гормон) стимулирует рост в яичнике везикулярного фолликула, секрецию фолликулярной жидкости, формирование оболочек, окружающих фолликул. У мужчин под его влиянием происходит образование сперматозоидов.
лютеинизирующий гормон (лютропин), необходим для роста везикулярного фолликула яичника на стадии, предществующей овуляции и для самой овуляции, т.е. разрыва оболочки созревшего фолликула и выхода из него яйцеклетки. Он стимулирует образование женских половых гормонов – эстрогенов. Под его воздействием происходит образование желтого тела из лопнувшего фолликула и стимуляция выработки им прогестерона. Лютропин способствует выработке андрогенов у мужчин.
Меланоцитстимулирующий гормон вырабатывается в средней доле гипофиза. Увеличивает содержание меланина в эпидермисе.
Липотропины (средняя доля) стимулируют освобождение жирных кислот из депо.
Гормоны нейрогипофиза
Задняя доля гипофиза тесно связана с супраоптическим и паравентрикулярным ядром гипоталамуса. Нервные клетки этих ядер вырабатывают нейросекрет, который транспортируется в заднюю долю гипофиза. Накапливаются гормоны в питуицитах, в этих клетках гормоны превращаются в активную форму. В нервных клетках паравентрикулярного ядра образуется окситоцин, в нейронах супраоптического ядра – вазопрессин.
Вазопрессин выполняет две функции:
1) усиливает сокращение гладких мышц сосудов (тонус артериол повышается с последующим повышением артериального давления);
2) угнетает образование мочи в почках (антидиуретическое действие). Антидиуретическое действие обеспечивается способностью вазопрессина усиливать обратное всасывание воды из канальцев почек в кровь. Уменьшение образования вазопрессина является причиной возникновения несахарного диабета (несахарного мочеизнурения).
Окситоцин избирательно действует на гладкую мускулатуру матки, усиливает ее сокращение. Сокращение матки резко увеличивается, если она находилась под воздействием эстрогенов. Во время беременности окситоцин не влияет на сократительную способность матки, так как гормон желтого тела прогестерон делает ее нечувствительной ко всем раздражителям. Окситоцин стимулирует выделение молока, усиливается именно выделительная функция, а не его секреция. Особые клетки молочной железы избирательно реагируют на окситоцин. Акт сосания рефлекторно способствует выделению окситоцина из нейрогипофиза.
Функциональная классификация гормонов
Рилизинг-гормоны – гормоны, регулирующие синтез и выделение гормонов аденогипофиза, преимущественно тропных. Синтезируются и выделяются нейросекреторными клетками гипоталамуса.
К известным рилизинг-гормонам относятся:
кортикотропин-рилизинг-гормон
соматотропин-рилизинг-гормон
тиреотропин-рилизинг-гормон
гонадотропин-рилизинг-гормон.
2. Тропные гормоны – гормоны, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Образуются и выделяются аденогипофизом. К тропным гормонам относятся:
адренокортикотропный гормон
тиреотропный гормон
фолликулостимулирующий гормон
лютеинизирующий гормон
соматотропный гормон
меланоцитстимулирующий гормон
3. Эффекторные гормоны – гормоны, которые оказывают влияние непосредственно на орган-мишень. Образуются и выделяются в основном периферическими железами внутренней секреции, а также гипофизом.
+ Йодированные гормоны щитовидной железы и их физиологическое значение. Физиологическое значение нейодированного гормона щитовидной железы. Гипофункция щитовидной железы. Гиперфункция железы. Секреция гормонов щитовидной железы.
Гормоны щитовидной железы делятся на две группы:
1) йодированные – тироксин, трийодтиронин;
2) тиреокальцитонин (кальцитонин).
Функции йодированных гормонов:
1. У растущего организма они участвуют в дифференцировке развивающихся клеток, влияя на процессы роста, физическое, умственное и половое развитие. Особенно важную роль эти гормоны играют в дифференцировке нервной системы у плода.
2. Участвуют в созревании и развитии нервной системы. Стимулируют в нейронах и глиоцитах экспрессию генов, кодирующих наиболее важные для нервной ткани белки (тубулины, основной белок миелина, синаптосины, фактор роста нервов и др.). Стимулируют развитие мозга и психическое развитие (формирование синапсов, миелинизацию аксонов, транспорт аминокислот через гематоэнцефалический барьер). Повышают возбудимость ЦНС и особенно симпатического отдела вегетативной нервной системы. Активируют гипоталамические центры, повышают аппетит и потребление пищи.
3. Усиливают все виды обмена веществ – белковый, углеводный, липидный. Синтез белков, в том числе и тканеспецифических, увеличивается в результате активации транскрипции в клетках мишенях. Йодтиронины являются синергистами гормона роста. Очень высокие концентрации гормонов, напротив, тормозят синтез белков и стимулируют катаболические процессы. Стимулируют все этапы метаболизма углеводов; ускоряют синтез холестерина и желчных кислот в печени, усиливают липолиз и окисление жирных кислот.
4. Повышают основной обмен и усиливают энергообразование в организме, при этом возрастает потребление О2 во всех тканях, кроме мозга, половых желез и селезенки. Повышают температуру тела.
5. Усиливают частоту сердечных сокращений (за счет увеличения количества β1-адренорецепторов), сердечный выброс, систолическое и пульсовое артериальные давления, кровоток в большинстве тканей, уменьшают сосудистый тонус.
6. Стимулируют деятельность органов пищеварительного тракта: усиливают моторику и увеличивают секрецию пищеварительных соков.
7. Усиливают эритропоэз, активируя выработку эритропоэтинов, а также увеличивая образование β2-адренорецепторов на плазмолемме эритроидных клеток.
8. Способствуют нормальной функции репродуктивной системы: стимулируют выработку гонадолиберина, ЛГ, тестостерона; увеличивают чувствительность яичников к гонадотропным гормонам, а эндометрия – к эстрогенам.
Секреция гормонов щитовидной железы.
1) образование коллоида, синтез тиреоглобулина;
2) йодирование коллоида (тиреоглобулина), поступление йода в организм, всасывание в виде йодидов. Йодиды поглощаются щитовидной железой, окисляются в элементарный йод и включаются в состав тиреоглобулина, процесс стимулируется ферментом – тиреоидпероксидазой;
3) выделение в кровоток происходит после гидролиза тиреоглобулина под действием катепсина. При этом освобождаются активные гормоны – тироксин, трийодтиронин (неактивные – монойодтирозин и дийодтирозин).
Тиреокальцитонин. Образуется парафолликулярными клетками щитовидной железы, которые расположены вне железистых фолликул. Он принимает участие в регуляции кальциевого обмена, под его влиянием уровень Ca снижается. Тиреокальцитоцин понижает содержание фосфатов в периферической крови. Кальцитонин участвует в регуляции обмена Са2+:
угнетение функции остеокластов и активация функции остеобластов, усиление процессов минерализации, угнетение реабсорбции Са2+ в почках и увеличение его выделения с мочой, гипокальциемия.
Секреции тиреокальцитонина способствуют некоторые биологически активные вещества: гастрин, глюкагон, холецистокинин.
При возбуждении бетаадренорецепторов повышается секреция гормона, и наоборот.
Недостаточность выработки гормона (гипотериоз), появляющаяся в детском возрасте, ведет к развитию кретинизма (задерживаются рост, половое развитие, развитие психики, наблюдается нарушение пропорций тела). Недостаточность выработки гормона ведет к развитию микседемы, которая характеризуется резким расстройством процессов возбуждения и торможения в ЦНС, психической заторможенностью, снижением интеллекта, вялостью, сонливостью, нарушением половых функций, угнетением всех видов обмена веществ.
При повышении активности щитовидной железы (гипертиреозе) возникает заболевание тиреотоксикоз. Характерные признаки: увеличение размеров щитовидной железы, числа сердечных сокращений, повышение обмена веществ, температуры тела, увеличение потребления пищи, пучеглазие. Наблюдаются повышенная возбудимость и раздражительность, изменяется соотношение тонуса отделов вегетативной нервной системы: преобладает возбуждение симпатического отдела. Отмечаются мышечное дрожание и мышечная слабость.
Недостаток в воде йода приводит к снижению функции щитовидной железы со значительным разрастанием ее ткани и образованием зоба. Разрастание ткани – компенсаторный механизм в ответ на снижение содержания йодированных гормонов в крови.
+ Гормоны коры надпочечников и их значение. Роль гормонов мозгового вещества надпочечников. Регуляция продукции альдостерона. Регуляция продукции глюкокортикоидов. Синтез катехоламинов.
Надпочечники – парные железы, расположенные над верхними полюсами почек. Они имеют важное жизненное значение. Различают два типа гормонов: гормоны коркового слоя и гормоны мозгового слоя.
Гормоны коркового слоя делятся на три группы:
1) глюкокортикоиды (гидрокортизон, кортизон, кортикостерон);
2) минералокортикоиды (альдестерон, дезоксикортикостерон);
3) половые гормоны (андрогены, эстрогены, прогестерон).