

20. Строение эпикарда.
Эпикард и перикард (имеют сходный структурный состав: мезотелий, РВСТ, жировая ткань, сосуды, нервный аппарат. Между ними находится перикардиальная полость В сердце имеется также фиброзный каркас (фиброзные кольца, треугольники, мембраны). Эти структуры построены из ПВСТ (может встречаться хрящевая ткань).
Эпикард покрыт мезотелием, под которым располагается рыхлая волокнистая соединительная ткань, содержащая сосуды и нервы. В эпикарде может иметься в значительном количестве жировая ткань. Эпикард представляет собой висцеральный листок перикарда; париетальный листок также имеет строение серозной оболочки и обратен к висцеральному слоем мезотелия. Гладкие влажные поверхности париетального и висцерального листков легко скользят друг по другу при сокращении сердца. При повреждении мезотелия (обычно вследствие
воспалительного процесса - перикардита) за счет образующихся между листками соединительнотканных спаек деятельность сердца может существенно нарушаться.
ОРГАНЫ ЭНДОКРИННОГО АППАРАТА
ПРЕПАРАТЫ
ГИПОФИЗ

Передняя доля гипофиза (фрагмент) (окраска гемалауном, большое увеличение):
1 - ацидофильная клетка; 2 - базофильная клетка (с макулой), гонадотропоцит; 3 - базофильная клетка (без макулы), тиротропоцит; 4 - хромофобные клетки; 5 - синусоидный капилляр
1 - ацидофильная клетка; 2 - базофильная клетка; 3 - хромофобная клетка; 4 - синусоидный капилляр


Уметь определять: 1 – передняя доля гипофиза; 1.1 – хромофобные клетки; 1.2 – хромофильные клетки; 1.2.1 – ацидофильный аденоцит; 1.2.2 – базофильный аденоцит; 1.2 – рыхлая соединительная ткань; 1.3 – синусоидные капилляры; 2 – задняя доля гипофиза; 2.1 – питуициты; 2.2 – отростки нейросекреторных клеток; 2.2.1 – тельца Херринга; 2.3 – фенестрированные капилляры.

Некоторые авторы промежуточную и переднюю часть объединяют в одно целое под названием «аденогипофиз».
Аденогипофиз и его гормоны
Гормоны передней доли гипофиза большей частью являются регуляторными. Это означает, что они координируют деятельность периферических эндокринных желез (их также называют «тропными» гормонами).
АКТГ
Адренокортикотропный гормон гипофиза (сокращенно – АКТГ) – главный стимулятор коры надпочечников. Он вызывает разрастание (гипертрофию) пучкового слоя органа и усиливает синтез в нем глюкокортикоидов (основных гормонов, обеспечивающих ответную реакцию на стресс и процессы адаптации).
АКТГ также проявляет меланоцитстимулирующую функцию, вызывает образование пигмента меланина (результат – пигментация кожи).
Гонадотропные гормоны
Лютеинизирующий гормон (ЛГ) и фолликулостимулирующий (ФСГ) гормон-вещества, отвечающие за репродуктивную систему человека. Это так называемые гонадотропные гормоны гипофиза. ЛГ стимулирует овуляцию и выработку эстрогенов у женщин, а также производство андрогенов у мужчин. Функция ФСГ: помощь в созревании фолликулов в яичниках и участие в сперматогенезе.
Тиреотропины
Тиреотропный гормон (сокращенно ТТГ) – главный координатор синтеза и секреции основных гормонов щитовидной железы (тироксина, трийодтиронина). Известно, что под влиянием ТТГ происходит увеличение количества и размеров клеток щитовидной железы. ТТГ также влияет на синтез нуклеотидов и фосфолипидов. При недостатке или избытке ТТГ у человека появляются проблемы со щитовидной железой (повышенная или пониженная функция органа).
СТГ
Соматотропин (СТГ) – гормон, главная функция которого – рост организма и синтез белка в клетках (так называемые анаболические процессы). Соматотропин также влияет на образование глюкозы и распад жиров. Именно соматотропин отвечает за рост тела и физическое развитие человека. Некоторые свои эффекты гормон соматотропин осуществляет опосредованно через печень, а также – через вилочковую железу.
Если соматотропин повышен в детском возрасте, это приводит к усиленному росту тела – гигантизму, более всего в длину увеличиваются конечности. Если же соматотропин повышается у взрослого человека, в размерах увеличиваются отдельные части тела – нос, губы, скулы, кисти и стопы. Заболевание называется акромегалией и может иметь наследственную предрасположенность. Соматотропин иногда повышается вследствие доброкачественной опухоли (аденомы) гипофиза.
Когда соматотропин снижается в детском возрасте, это заканчивается низкорослостью человека. В медицине данное состояние называется нанизмом или карликовым ростом. Причиной нанизма может быть врожденное недоразвитие передней части гипофиза или разрушение этого участка опухолью.
Пролактин
Гормон пролактин регулирует выделение молока у млекопитающих, а также оказывает и другие функции:
пролактин влияет на дифференцировку различных тканей;
воздействует на рост и обмен веществ;
пролактин обеспечивает инстинкт выхаживания потомства у млекопитающих;
пролактин способствует появлению молока у кормящей женщины и стимулирует рост грудных желез;
пролактин содействует переходу молозива в женское молоко;
принимает участие в формировании вторичных половых признаков у женщины;
пролактин у мужчин влияет на рост простаты;
в период выкармливания ребенка грудным молоком пролактин предупреждает наступление месячных и новое зачатие плода.
Низкий уровень пролактина приводит к нарушению менструального цикла у женщин и половой дисфункции у мужчин.
Средняя доля
В этом участке гипофиза вырабатывается меланотропин, который влияет на пигментацию эпителия. Предполагается, что меланотропин участвует в формирование памяти.
Гормоны задней части
К ним относятся окситоцин и вазопрессин, вырабатываемые в ядрах гипоталамуса. Гипофиз для них выполняет функцию резервуара. Окситоцин влияет на сократительную функцию матки, повышает количество пролактина, активизирует выделение молозива у женщины.
Вазопрессин повышает реабсорбцию воды в почечных канальцах и уменьшает выделение мочи. Другая функция вазопрессина – стимулирующее воздействие на гладкую мускулатуру (матки, кишечника, сосудов). В больших концентрациях вазопрессин повышает АД.
ЩИТОВИДНАЯ ЖЕЛЕЗА

1 – фолликул; 1.1 – тироцит; 1.2 – коллоид; 2 – интерстициальная соединительная ткань; 2.1 – кровеносные сосуды; 2.2 – интерфолликулярный островок.


ГИПОТАЛАМО-ГИПОФИЗАРНАЯ СИСТЕМА



НАДПОЧЕЧНИК
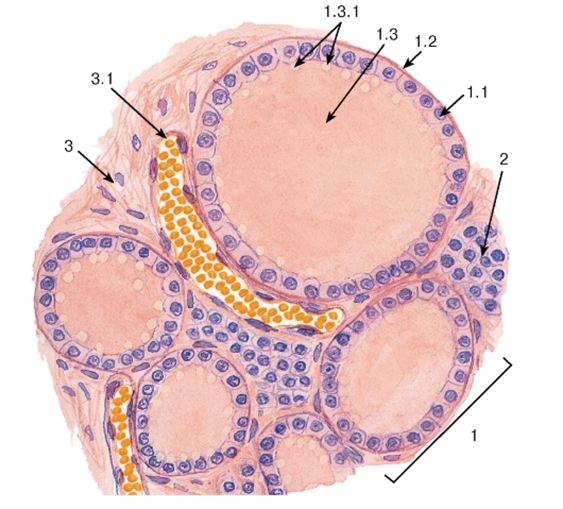
1 - соединительнотканная капсула; 2 - корковое вещество: А - клубочковая зона; Б - пучковая зона; В - сетчатая зона; Г - капилляры сетчатой зоны; 3 - мозговое вещество; 4 - хромаффинные клетки; 5 - кровеносные сосуды мозгового вещества

ТЕРМИНЫ
Аутокринная регуляция - При аутокринной регуляции клетка—продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка—продуцент гормона в то же время является его мишенью).
Аксо-вазальный синапс – (СИНАПС - это место передачи нервных импульсов с одной нервной клетки на другую) такая вот связь между аксоном и веной.
Аденогипофиз - (передняя доля, промежуточная доля, туберальная часть)
Ацидофилы - (около 40% всех аденоцитов) - мелкие округлые клетки с хорошо развитыми органеллами и высоким содержанием крупных гранул(аденоцит-клетка аденогипофиза)
Гипоталамус - небольшая область в промежуточном мозге, включающая в себя большое число групп клеток (свыше 30 ядер), которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма.
Гипофиз - это самый главный орган эндокринной системы. Анатомически он тесно связан с гипоталамусом, который через нейропептиды посылает гипофизу команды. Гипофиз выделяет гормоны, отвечающие за обмен веществ, рост и репродуктивную функцию.
Гонадотропы - подкласс тропных гормонов передней доли гипофиза и плаценты, физиологической функцией которых является регуляция работы половых желёз.
Главные паратироциты - гормонально-активные клетки, которые подразделяются на светлые клетки, преобладающие у детей, и темные клетки, преобладающие у взрослых.
Гормоны - биологически активные вещества органической природы, вырабатывающиеся в специализированных клетках желёз внутренней секреции (эндокринные железы), поступающие в кровь, связывающиеся с рецепторами клеток-мишеней и оказывающие регулирующее влияние на обмен веществ и физиологические функции.
Кортикотропы - это базофильные клетки в передней доле гипофиза, которые продуцируют проопиомеланокортин (ПОМК), который расщепляется на адренокортикотропин (АКТГ), β-липотропин (β-LPH) и меланоцит-стимулирующий гормон (MSH)
Коллоид - негормональное белковое вещество
Клубочковая зона (надпочечников) - слой эпителиальных клеток в корковом веществе надпочечника, расположенных в виде аркад или клубков и вырабатывающих минералокорти
Клетки диффузной эндокринной системы открытого типа - являются хеморецепторами содержимого полых органов, сообщающихся с внешней средой.
Хромаффинные клетки - нейроэндокринная клетка мозгового вещества надпочечников и параганглиев. Эмбриональным источником хромаффинных клеток служит ганглиозная пластинка.
Щитовидная железа - эндокринная железа у позвоночных, хранящая йод и вырабатывающая йодсодержащие гормоны (йодтиронины), участвующие в регуляции обмена веществ и росте отдельных клеток, а также организма в целом — тироксин (тетрайодтиронин, T4) и трийодтиронин (T3).
Эндокринная регуляция - система регуляции деятельности внутренних органов посредством гормонов, выделяемых эндокринными клетками непосредственно в кровь либо диффундирующих через межклеточное пространство в соседние клетки.
Эпифиз (или шишкови́дное те́ло) - эндокринная железа неврогенной группы, представлена небольшим шишковидным телом серовато-красноватого цвета, располагающимся в области четверохолмия среднего мозга.
Клетки диффузной эндокринной системы закрытого типа - Клетки закрытого типа не достигают полости органа и получают информацию о состоянии внутренней среды организма
Лактотропы , или клетки, продуцирующие пролактин – гормона, отвечающий за образование молока в молочных железах, составляют около 1\5 всех клеток передней доли гипофиза и располагаются в заднебоковых отделах.
Мелкоклеточные ядра гипоталамуса - вырабатывают ряд гипофизотропных факторов, которые усиливают (рилизинг факторы или либерины) или угнетают (ингибирующие факторы или статины) выработку гормонов клетками передней доли, попадая к ним по воротной системе сосудов.
Надпочечники - парные эндокринные железы, расположенные над верхней частью почек позвоночных животных и человека. У человека расположены в непосредственной близости к верхнему полюсу каждой почки.
Нейроэндокринная регуляция - дна из форм физиологической регуляции в организме человека и животных, при которой нервные импульсы и переносимые кровью и лимфой вещества (метаболиты, гормоны, а также другие нейромедиаторы) принимают совместное участие в едином регуляторном процессе.
Нейросекреторные клетки - Нервная и эндокринная системы, представляя собой единый комплекс, имеют и свой морфологический субстрат — нейросекреторные клетки, которые занимают промежуточное положение между нервными и эндокринными элементами. Совмещая нервную и эндокринную функции, нейросекреторные клетки с одной стороны воспринимают импульсы, поступающие к ним из других отделов нервной системы, а с другой — передают эти импульсы в кровь посредством нейрогормонов.
Нейрогипофиз - анатомически подразделяется на заднюю часть или долю и гипофизарную ножку, связывающую гипофиз с мозгом. В Н. поступают нервные волокна нейросекреторных ядер гипоталамуса.
Околощитовидные железы - это две пары маленьких желёз, которые расположены по бокам от щитовидной железы на шее. Заболевание проявляется повышенной продукцией паратгормона и различными клиническими проявлениями, к которым чаще всего относятся повышение кальция в крови, остеопороз, мочекаменная болезнь и язвы желудка.
Оксифильные паратироциты - появляются в возрасте 10 лет, они гормонально-неактивны. Основным гормоном ПЩЖ является паратгормон, который образуется в околощитовидных железах из предшественников препропаратгормона и пропаратгормона.
Паракринная регуляция - вариант воздействия гормона на клетку - мишень. При паракринной регуляции продуцент биологически активного вещества - гормона и клетка мишень расположены рядом. Молекулы гормона достигают мишени посредством диффузии в межклеточном веществе.
Паравентрикулярное ядро - группа нейронов гипоталамуса.
Питуициты - мелкие глиальные клетки с многочисленными отростками, образующие строму задней доли. (ЭТО ПО ГИПОФИЗУ)
Пинеалоциты - это основные клетки, содержащиеся в шишковидной железе , расположенной за третьим желудочком и между двумя полушариями мозга . Основная функция пинеалоцитов - секреция гормона мелатонина , важного для регуляции циркадных ритмов .
Пучковая зона - В пучковой зоне образуются глюкокортикоиды, к которым относятся:
• Кортизол
• Кортизон
(ОТНОСИТСЯ К НАДПОЧЕЧНИКАМ)
Супраоптическое ядро - у человека содержит в себе около 3 000 нейронов. Они синтезируют гормон вазопрессин, который по кровотоку достигает сосочковых протоков почек и повышает реабсорбцию (обратное всасывание) воды. (ОТНОСИТСЯ К ГИПОТАЛАМУСУ)
Соматотропы - вырабатывают соматотропный гормон (СТГ) или
гормон роста (ГР); его эффект стимуляции роста опосредован особыми пептидами - соматомединами
С-клетки - эндокринные клетки слизистой оболочки тонкой кишки, секретирующие секретин. S-клетки относятся к апудоцитам и входят в состав гастроэнтеропанкреатической эндокринной системы, которая является частью диффузной эндокринной системы (синоним АПУД-система).
ТО ВОТ Ц - C-клетки (парафолликулярные), относящиеся к диффузной эндокринной системе, секретируют кальцитонин — один из факторов, регулирующих обмен кальция в клетках, участник процессов роста и развития костного аппарата (наряду с другими гормонами).
Сетчатая зона - В сетчатой зоне производятся половые гормоны (андрогены, являющиеся веществами — предшественниками эстрогенов). Данные половые гормоны играют роль несколько иную, чем гормоны, выделяемые половыми железами.
Стероидогенез - биологический процесс, при котором стероиды образуются из холестерина и превращаются в другие стероиды.
Тельца Херринга - глыбки нейросекрета в задней доле гипофиза, сместившиеся из переднего гипоталамуса по аксонам гипоталамо гипофизарного пучка
Тиротропы - вырабатывают тиротропный гормон (ТТГ), который усиливает активность тироцитов.
Тироциты - Тироксин (Т4) и трийодтиронин (Т3) регулируют обмен веществ, рост и развитие организма. Синтез этих гормонов происходит в эпителиальных фолликулярных клетках, называемых тироцитами
Фолликулярно-звездчатые клетки - несекреторные, звездчатой формы,
охватывающие своими отростками секреторные клетки и выстилающие мелкие фолликулярные структуры. Способны фагоцитировать гибнущие клетки и
влиять на секреторную активность базофилов и ацидофнлов.
Фолликул щитовидной железы - клетки, расположенные в щитовидной железе и способные к синтезу и секреции тиреоидных гормонов, таких как тироксин и трийодтиронин.
Хромофильные аденоциты - аденоциты, хорошо воспринимающие гистологические красители.
Хромофобные аденоциты - слабо окрашивающиеся А., не содержащие секреторных гранул.
ВОПРОСЫ
Общая характеристика органов эндокринного аппарата
Эндокринными называют железы, продукты которых (гормоны) выделяются непосредственно в кровь.
Гормоны - вещества с высокой биологической активностью - регулируют рост и деятельность клеток различных тканей (клеток-мишеней) благодаря наличию на последних специфических рецепторов гормонов.
Эндокринная регуляция является одним из нескольких известных видов регуляторных воздействий (рис. 3-1), среди которых выделяют:

(1)аутокринное (в пределах клеток одного типа);
(2)паракринное (воздействие продуктов клеток одного типа на клетки другого типа);
(3)эндокринное (опосредованное гормонами, циркулирующими в крови);
(4)нервное (воздействие нейрона на орган-мишень через отросток, опосредованное ножным скончанием);
(5)нейроэндокринное (сочетает признаки эндокринного и нервного).
Воздействия (1) и (2) являются локальными, (3), (4) и (5) - дистантными.
Строение эндокринных желез достаточно разнообразно. Они могут быть одноклеточными (элементы ДЭС - диффузной эндокринной системы), иметь вид мелких компактных клеточных скоплений (панкреатические островки) или являться сравнительно крупными органными структурами (щитовидная и околощитовидная железы, эпифиз, гипофиз, надпочечник). Некоторые эндокринные железы (гипофиз, надпочечник) образованы тканями, имеющими различное эмбриональное происхождение и у низших позвоночных расположенными раздельно. Другие (островки Лангерганса, эндокринные элементы кишечной трубки, половых желез) - лежат внутри неэндокринных. Более тело, есть эндокринные органы, периодически появляющиеся и исчезающие (желтое тело и фолликул яичника).
Эндокринные железы, имеющие органное строение, обычно покрыты капсулой из плотной соединительной ткани, от которой вглубь отходят истончающиеся трабекулы, состоящие из рыхлой соединительной ткани и несущие сосуды и нервы. В большинстве эндокринных желез клетки образуют тяжи и тесно прилежат к капиллярам, что обеспечивает секрецию гормонов в кровоток. Капилляры формируют очень густые сети и благодаря своему строению обладают повышенной проницаемостью - они являются фенестрированными или синусоидными. Так как гормоны выделяются в кровь,выводные протоки у желез отсутствуют.
Клетки эндокринных желез характеризуются высокой секреторной активностью и значительным развитием синтетического аппарата; их строение зависит, в первую очередь, от химической природы вырабатываемых гормонов. В клетках, образующих пептидные гормоны, сильно развиты грЭПС, комплекс Гольджи; в синтезирующих стероидные гормоны, - аЭПС, митохондрии с тубулярно-везикулярными кристами.
Доказана способность многих клеток одновременно вырабатывать несколько гормонов; это позволило отвергнуть ранее сформулированный принцип "одна клетка - один гормон". Полиептидные гормоны нередко синтезируются в виде крупных молекул-предшественников, которые в дальнейшем подвергаются внутриклеточной ферментной обработке (процессингу) с отщеплением активных гормонов. Различные типы клеток (например, в гипофизе) могут исходно синтезировать один вид молекулыпредшественника, однако вследствие специфического процессинга из нее образуются разные гормоны.
Накопление гормонов обычно происходит внутриклеточно в виде секреторных гранул; нейрогормоны могут аккумулироваться в больших количествах внутри аксонов, резко растягивая их в отдельных участках
(накопительные нейросекреторные тельца). Единственный пример внеклеточного накопления гормонов - в фолликулах щитовидной железы.
Функционально ведущая ткань эндокринной железы - клетки, которые продуцируют гормоны. Способностью к эндокринной секреции обладают все труппы тканей, имеющиеся в организме.
Эпителий - функционально ведущая ткань большинства эндокринных желез (щитовидная и околощитовидные железы, передняя и промежуточная доли гипофиза, корковое вещество надпочечника, некоторые эндокринные элементы гонад - фолликулярные клетки яичника, сустентоциты яичка и др.). Он может относиться к различным гистогенетическим типам.
Соединительнотканное происхождение имеет часть эндокринных элементов гонад (интерстициальные эцдокриноциты - клетки Лейдига, клетки внутреннего слоя теки фолликулов яичника и др.).
Нейральное происхождение имеют эндокринные клетки гипоталамуса, эпифиза, нейрогипофиза, мозгового вещества надпочечнику, элементы дэс.
К мышечным тканям относятся юкстагломерулярные клетки в средней оболочке артериол почечного клубочка, а также секреторные кардиомиоциты.
Общие закономерности организации эндокринной системы включают:
иерархический принцип и (2) наличие системы обратных связей.
1. Иерархический принцип демонстрируется наличием в эндокринной системе нескольких уровней организации. Нижний из них занимают железы, вырабатывающие гормоны, которые влияют на различные ткани организма
(эффекторные или периферические). Деятельность большинства этих желез регулируется особыми тропными гормонами передней доли гипофиза (второй, более высокий уровень). В сдою очередь, выделение тронных гормонов контролируется специальными нейрогормонами гипоталамуса, который и занимает наиболее высокое положение в иерархической организации системы.
В соответствии с иерархическим принципом, нарушение деятельности периферической железы может обусловливаться изменениями как в самой железе, так и в выделении соответствующих гипофизарных или гипоталамических факторов. Выявление уровня повреждения имеет существенное клиническое значение. В более редких случаях отсутствие эффекта гормонов связано с дефектом соответствующих рецепторов на тканяхмишенях (как правило, генетически обусловленным).
2. Система обратных связей (обычно отрицательных) обеспечивает поддержание необходимого уровня активности эндокринных желез вследствие того, что усиление выработки гормонов периферическими железами угнетает, а ослабление - стимулирует секрецию соответствующих тропных гормонов гипофиза и факторов гипоталамуса.
Эндокринная система взаимодействует с другими регуляторными системами, в частности, иммунной и нервной. Благодаря иннервации эндокринные железы контролируются нервной системой, на деятельность которой, в свою очередь, оказывают влияние гормоны. Особенно ярко взаимосвязь нервной и эндокринной систем проявляется в том, что центральный орган эндокринной системы, интегрирующий ее функцию с деятельностью других систем организма - гипоталамус - сам является частью ЦНС.
Эндокринная система включает:
центральное звено: гипоталамус, гипофиз, эпифиз
Функция: регуляция работы эндокринных желез.
периферическое звено: эндокринные железы и эндокринные клетки.
Функция: регуляция работы организма.
Эндокринную систему разделяют на гландулярную (железы внутренней секреции) и
диффузную.
Эндокринные железы:
гипофиз (аденогипофиз и нейрогипофиз);
щитовидная железа;
околощитовидные (паращитовидные) железы;
надпочечники;
эпифиз
Органы с эндокринной тканью:
поджелудочная железа (островки Лангерганса);
половые железы (семенники и яичники)
Органы с эндокринными клетками:
ЦНС (в особенности — гипоталамус);
сердце;
легкие;
желудочно-кишечный тракт;
почка;
плацента;
тимус
предстательная железа
Железы внутренней секреции осуществляют выработку гормонов, к которым
причисляются все стероидные гормоны, а также гормоны щитовидной железы и
некоторые пептидные гормоны (гормоны, состоящие из аминокислот). Диффузную
эндокринную систему представляют рассеянные по всему телу эндокринные клетки,
которые производят пептидные гормоны.
Практически любая ткань организма содержит эндокринные клетки.
Функции эндокринной системы:
1. гуморальная регуляция функций организма;
2. координация работы всех органов и систем;
3. гомеостаз организма при изменяющихся условиях внешней среды;
4. рост и развитие организма;
5. половая дифференцировка и репродуктивная функция;
6. обмен веществ и энергии;
7. эмоциональные реакции;
8. психическая деятельность человека.
Характеристика ядер гипоталамуса.
Нейросекреторные ядра гипоталамуса в зависимости от размеров клеток и их функциональных особенностей разделяют на крупно- и мелкоклеточные.
1. Крупноклеточные ядра образованы клеточными телами, которые в 2-3 раза крупнее, чем в других отделах гипоталамуса: к ним относятся супраоптическое (СОЯ) и паравентрикулярное (ПВЯ) ядра. СОЯ в 3-4 раза объемнее ПВЯ: ПВЯ лишь в центральных участках построено по типу крупноклеточного, а в периферических отделах образовано мелкими нейросекреторными клетками. Аксоны клеток СОЯ и ПВЯ покидают гипоталамус и, в составе гипоталамо-гипофизарного тракта пересекая гемато-энцефалический барьер, проникают в заднюю долю гипофиза, где образуют терминали на капиллярах (рис. 3-3).
Крупноклеточные ядра секретируют антндиуретическнй гормон (АДГ) или вазопрессин (ВП), и окситоцин. Эти гормоны вырабатываются разными клетками. У человека АДГ образуется, главным образом, в СОЯ, а окситоцин - в ПВЯ. АДГ и окситоцин синтезируются в грЭПС (в виде крупной молекулы прогормона) и переносятся в комплекс Гольджи, где упаковываются в гранулы. Процессинг продукта (завершается лишь в ходе транспорта гранул в аксоне) приводит к освобождению активного гормона и нейрофизина - белка с неясной функцией (ранее считали переносчиком нейрогормонов).
2. Мелкоклеточные ядра вырабатывают ряд гипофизотропных факторов, которые усиливают (рилизинг факторы или либерины) или угнетают (ингибирующие факторы или статины) выработку гормонов клетками передней доли, попадая к ним по воротной системе сосудов. Аксоны нейросекреторных клеток этих ядер образуют терминали на первичной капиллярной сети в срединном возвышении (нейрогемальной контактной зоне). Эта сеть далее собирается в воротные вены, проникающие в переднюю долю гипофиза и
распадающиеся на вторичную сеть капилляров между тяжами железистых клеток аденоцитов (см. рис. 3-3).
Строение нейросекреторных клеток.
Нейросекреторные клетки - отростчатой формы, с крупным везикулярным ядром, хорошо заметным ядрышком и базофильной цитоплазмой, содержащей развитую грЭПС и крупный комплекс Гольджи, от которого отделяются нейросекреторные гранулы (рис. 3-2). Гранулы транспортируются со скоростью около 1-4 мм/ч по аксону вдоль центрального пучка микротрубочек и микрофиламентов, а местами накапливаются в больших количествах, растягивая аксон. Самые крупные из таких участков хорошо видны под световым микроскопом и называются накопительными нейросекреторными тельцами (Херринга): в них сосредоточено до 60% всего нейросекрета, лишь около 30% находится в области терминалей. Терминали (аксо-вазальные синапсы) характеризуются присутствием, помимо гранул, многочисленных светлых пузырьков (осуществляют возврат мембраны после экзоцитоза).
Понятие о гипоталамо-гипофизарной системе.
Это нейроэндокринный комплекс позвоночных, образованный гипоталамусом и гипофизом. Является осн. звеном в цепи нервной регуляции эндокринной функции организма. Информация, поступающая к нейронам гипоталамуса из внешней и внутр. среды организма в виде электрич. импульсов, трансформируется в них в химич. сигналы (нейрогормоны), поступающие в кровоток и оказывающие регуляторное влияние на гипофиз и периферич. органы-мишени.
У млекопитающих, исходя из особенностей организации функционирования и эмбрионального развития, выделяют две подсистемы: гипоталамо-нейрогипофизарную и гипоталамо-аденогипофизарную. Гипоталамо-нейрогипофизарная система образована крупными нейронами супраоптического и паравентрикулярного ядер гипоталамуса; их аксоны оканчиваются в задней доле гипофиза (нейрогипофизе) на высокопроницаемых капиллярах системы общего кровотока, доставляя туда гормоны вазопрессин и окситоцин. Осн. физиологич. сигналом, вызывающим выделение вазопрессина, является повышенная осмоляльность крови, а стимулом для секреции окситоцина служит сосание молока у лактирующей самки.
Гипоталамо-аденогипофизарная система образована связанными между собой мелкими нейронами, входящими в состав мн. ядер гипоталамуса, и разл. отделами передней доли гипофиза (аденогипофиза). Аксоны большинства нейронов гипоталамуса оканчиваются в т. н. срединном возвышении на высокопроницаемых капиллярах портальной системы циркуляции крови, связывающей гипоталамус с передней долей гипофиза. Секретируемые ими пептидные и моноаминовые нейрогормоны (дофамин, норадреналин, серотонин) с кровью поступают в аденогипофиз. Пептидные нейрогормоны оказывают стимулирующее (рилизинг-гормоны, или либерины) или ингибирующее (статины) влияние на выделение т. н. тропных гормонов, регулирующих деятельность периферич. эндокринных желёз. Напр., тиреолиберин стимулирует секрецию тиреотропина, который, в свою очередь, стимулирует активность щитовидной железы, вызывая секрецию в кровь её гормонов. У большинства млекопитающих дофаминергич. аксоны оканчиваются не только в срединном возвышении гипоталамуса, но и в промежуточной доле гипофиза на железистых клетках, выделяющих гормон меланотропин. Дофамин оказывает ингибирующее действие на секрецию этого гормона. У человека в ходе индивидуального развития происходит инволюция промежуточной доли гипофиза, поэтому у взрослых людей отсутствует её прямая нервная регуляция. В пределах гипоталамо-аденогипофизарной системы существует регуляция по принципу ультракороткой и короткой обратной связи. В первом случае сами нейрогормоны, а во втором случае гормоны аденогипофиза регулируют секреторную активность нейронов гипоталамуса.
Развитие и строение аденогипофиза. (передняя доля, промежуточная доля, туберальная часть)
Аденогипофиз представляет собой большую часть гипофиза и составляет около 75% от всей его массы. Он состоит из железистых клеток, которые, словно соты в улее разделены многочисленными тяжами-трабекулами.
Железистые клетки разделены на 5 основные видов по типу вырабатываемых ими гормональных веществ: соматотрофы, лактотрофы, кортикотрофы, тиротрофы, гонадотрофы.
аденогипофиз (передняя доля, промежуточная доля, туберальная часть)
Передняя доля гипофиза у человека составляет около 75% его массы; она образована анастомозирующими тяжами (трабекулами) аденоцитов тесно связанными с системой синусоидных капилляров. Форма аденоцитов варьирует от овальной до полигональной. На основании особенностей окраски их цитоплазмы выделяют: 1) хромофильные (интенсивно окрашивающиеся) и 2) хромофобные (слабо воспринимающие красители) клетки, которые содержатся примерно в равных количествах (рис. 3-5).
1. Хромофильные аденоциты (хромофилы) характеризуются развитым синтетическим аппаратом и накоплением в цитоплазме секреторных гранул, содержащих гормоны (рис. З-6). В зависимости от окраски секреторных гранул хромофилы подразделяют на ацидофилы и базофилы.
а) ацидофилы (около 40% всех аденоцитов) - мелкие округлые клетки с хорошо развитыми органеллами и высоким содержанием крупных гранул - включают два типа:
(1)соматотропы - вырабатывают соматотропный гормон (СТГ) или
гормон роста (ГР); его эффект стимуляции роста опосредован особыми пептидами - соматомединами;
(2)лактотропы - вырабатывают пролактин (ПРЛ) или лактотропный гормон (ЛТГ), который стимулирует развитие молочных желез и лактацию.
6) базофилы (10-20%) крупнее ацидофилов, однако их гранулы мельче и обычно содержатся в меньшем количестве. Включают гонадотропы,
тиротропы и адренокортикотропы:
(1)гонадотропы - вырабатывают а) фолликулостимулирующий гормон
(ФСГ), который стимулирует рост фолликулов яичника и сперматогенез, и б)
лютеинизирующий гормон (ЛГ), который способствует секреции женских и мужских половых гормонов, обеспечивает развитие овуляции и формирование желтого тела.
(2)тиротропы - вырабатывают тиротропный гормон (ТТГ), который усиливает активность тироцитов.
(3)кортикотропы - вырабатывают адренокортикотропный гормон
(АКТГ), который стимулирует активность коры надпочечника и является продуктом расщепления крупной молекулы проопиомеланокортина (ПОМК). ПОМК образует также МСГ и ЛПГ (см. ниже).
2. Хромофобные аденоциты (хромофобы) - разнородная группа клеток,
которая включает:
(1)хромофилы после выведения секреторных гранул,
(2)малодифференцированные камбиальные элементы, способные превращаться в базофилы или ацидофилы,
(3)фолликулярно-звездчатые клетки - несекреторные, звездчатой формы,
охватывающие своими отростками секреторные клетки и выстилающие мелкие фолликулярные структуры. Способны фагоцитировать гибнущие клетки и
влиять на секреторную активность базофилов и ацидофнлов.
Промежуточная доля у человека развита очень слабо и состоит из узких прерывистых тяжей базофильных и хромофобных клеток, которые секретируют
МСГ - меланоцитостимулирующий гормон (активирует меланоциты) и
ЛПГ - липотропный гормон (стимулирует обмен жиров). МСГ и ЛПГ (как и АКТГ) являются продуктами расщепления ПОМК. Встречаются кистозные полости, выстланные реснитчатыми клетками и содержащие негормональное белковое вещество - коллоид.
Туберальная часть в виде тонкого (25-60 мкм) рукава покрывает гипофизарную ножку, отделяясь от нее узким слоем соединительной ткани. Она состоит из тяжей хромофобных и хромофильных клеток;
Характеристика хромофобных аденоцитов.
Хромофобные аденоциты (хромофобы) - разнородная группа клеток,
которая включает:
(1)хромофилы после выведения секреторных гранул,
(2)малодифференцированные камбиальные элементы, способные превращаться в базофилы или ацидофилы,
(3)фолликулярно-звездчатые клетки - несекреторные, звездчатой формы,
охватывающие своими отростками секреторные клетки и выстилающие мелкие фолликулярные структуры. Способны фагоцитировать гибнущие клетки и
влиять на секреторную активность базофилов и ацидофнлов
Характеристика хромофильных аденоцитов.
Хромофильные аденоциты (хромофилы) характеризуются развитым синтетическим аппаратом и накоплением в цитоплазме секреторных гранул, содержащих гормоны (рис. З-6). В зависимости от окраски секреторных гранул хромофилы подразделяют на ацидофилы и базофилы.
а) ацидофилы (около 40% всех аденоцитов) - мелкие округлые клетки с хорошо развитыми органеллами и высоким содержанием крупных гранул - включают два типа:
(1)соматотропы - вырабатывают соматотропный гормон (СТГ) или
гормон роста (ГР); его эффект стимуляции роста опосредован особыми пептидами - соматомединами;
(2)лактотропы - вырабатывают пролактин (ПРЛ) или лактотропный гормон (ЛТГ), который стимулирует развитие молочных желез и лактацию.
6) базофилы (10-20%) крупнее ацидофилов, однако их гранулы мельче и обычно содержатся в меньшем количестве. Включают гонадотропы,
тиротропы и адренокортикотропы:
(1)гонадотропы - вырабатывают а) фолликулостимулирующий гормон
(ФСГ), который стимулирует рост фолликулов яичника и сперматогенез, и б)
лютеинизирующий гормон (ЛГ), который способствует секреции женских и мужских половых гормонов, обеспечивает развитие овуляции и формирование желтого тела.
(2)тиротропы - вырабатывают тиротропный гормон (ТТГ), который усиливает активность тироцитов.
(3)кортикотропы - вырабатывают адренокортикотропный гормон
(АКТГ), который стимулирует активность коры надпочечника и является продуктом расщепления крупной молекулы проопиомеланокортина (ПОМК). ПОМК образует также МСГ и ЛПГ
Регуляция деятельности клеток передней доли гипофиза.
Передняя доля сокрыта соединительной капсулой, от которой отходят прослойки соединительной ткани, которые составляют строму органа. Паренхимой органа являются эпителиальные клетки аденоциты, которые укомплектовываются в тяжи.
Клетки передней доли:
1. хромофильные (содержат гранулы, которые окрашиваются красителями)
• базофильные (10%)
-гонадотропные
-тиротропные
• ацидофильные
-соматотропные
-мамматропные
2. хромофобные (гранул не содержат, поэтому не окрашиваются) (60%)
• недифференцированные
• дифференцирующиеся
• хромофильные зрелые
• звездчато-фолликулярные
3. кортикотропные
Гонадотропные эндокриноциты — наиболее крупные клетки, имеют круглую, иногда угловатую форму, овальное или круглое ядро, смещенное к периферии, так как в центре клетки находится макула (пятно), в которой располагаются комплекс Гольджи и клеточный центр. В цитоплазме хорошо развиты гранулярная ЭПС, митохондрии и комплекс Голь¬джи, а также базофильные гранулы диаметром 200-300 нм, состоящие из гликопротеидов и окрашивающиеся альдегид-фуксином. Полагают, что существуют 2 разновидности гонадотропных эндокриноцитов, одни из которых выделяют фоллитро¬пин, другие — лютропин.
Фолликулотропный гормон (фоллитропин) в мужском ор¬ганизме действует на начальный этап сперматогенеза, в женском — на рост фолликулов и выделение эстрогенов в половых железах.
Лютропин стимулирует секрецию тестостерона в муж¬ских половых железах и развитие и функцию желтого тела в женских половых железах.
Клетки кастрации появляются в передней доле в тех слу¬чаях, когда половые железы вырабатывают недостаточное количество половых гормонов.
Тиротропные эндокриноциты имеют овальную или вы¬тянутую форму, овальное ядро. В их цитоплазме хорошо ра¬звиты комплекс Гольджи, гранулярная ЭПС и митохондрии, содержатся базофильные гранулы размером 80-150 нм, окрашивающиеся альдегидфуксином. Тиротропные эндо¬криноциты под влиянием тиролиберина вырабатывают тиротропный гормон, который стимулирует выделение тирок¬сина щитовидной железой.
Клетки тироидэктомии появляются в гипофизе при понижении функции щитовидной железы. В этих клетках ги¬пертрофируется гранулярная ЭПС, расширяются ее цистер¬ны, повышается секреция тиротропного гормона. В результа¬те расширения канальцев и цистерн ЭПС цитоплазма клеток приобретает ячеистый вид.
Кортикотропные эндокриноциты не относятся ни к аци¬дофильным, ни к базофильным, имеют неправильную форму, дольчатое ядро, в их цитоплазме содержатся мелкие гранулы. Под влиянием кортиколиберинов, вырабатываемых в ядрах медиобазального гипоталамуса, эти клетки секретируют кортикотропный или адренокортикотропный гормон (АКТГ), стимулирующий функцию коры надпочечников.
Ацидофильные эндокриноциты составляют 35-40 % и подразделяются на 2 разновидности, которые име¬ют обычно круглую форму, овальное или круглое ядро, распо¬ложенное в центре. В клетках хорошо развит синтетический аппарат, т. е. комплекс Гольджи, гранулярная ЭПС, митохон¬дрии; в цитоплазме содержатся ацидофильные гранулы.
Соматотропные эндокриноциты содержат гранулы овальной или круглой формы диаметром 400-500 нм, выра¬батывают соматотропный гормон, который стимулирует рост тела в детском и юношеском возрасте. При гиперфунк¬ции соматотропных клеток после завершения роста развива¬ется акромегалия — заболевание, характеризующееся появлением горба, увеличением размеров языка, нижней че¬люсти, кистей рук и стоп ног.
Маммотропные эндокриноциты содержат удлиненные гранулы, достигающие размеров 500-600 нм у рожениц и бе¬ременных женщин. У некормящих матерей гранулы умень¬шаются до 200 нм. Эти аденоциты выделяют маммотропный гормон, или пролактин. Функции: 1) стимулирует синтез мо¬лока в молочных железах; 2) стимулирует развитие желтого тела в яичниках и секрецию прогестерона.
Хромофобные (главные) эндокриноциты составляют око¬ло 60 %, имеют меньшие размеры, не содержат окрашивае¬мых гранул, поэтому их цитоплазма не окрашивается. В со¬став хромофобных аденоцитов входит 4 группы:
1) недиффе¬ренцированные (выполняют регенераторную функцию);
2) дифференцирующиеся, т. е. начали дифференцироваться, но дифференцировка не закончилась, в цитоплазме появи¬лись лишь единичные гранулы, поэтому цитоплазма слабо окрашивается;
3) хромофильные зрелые клетки, которые только что выделили свои секреторные гранулы, поэтому уменьшились в размере, а цитоплазма утратила способность к окрашиванию;
4) звездчато-фолликулярные клетки, харак¬теризующиеся длинными отростками, распространяющи¬мися между эндокриноцитами.
Группа таких клеток, обращенных апикальными поверхностями друг к другу, выделяет секрет, в результате чего образуются псевдофолликулы, за¬полненные коллоидом.
Задняя доля содержит:
(1)отростки и терминала нейросекреторных клеток СОЯ и ПНЯ гипоталамуса, по которым транспортируются и выделяются в кровь АДГ и окситоцин; расширенные участки по ходу отростков и в области терминалей называются накопительными нейросекреторными тельцами (Херринга);
(2)многочисленные фенестрированные капилляры;
(3)питуициты - отростчатые глиальные клетки (занимают до 25-30% объема доли) - образуют 3х-мерные сети, охватывают аксоны и терминалы нейросекреторных клеток и выполняют поддерживающую и трофическую функции, а также, возможно, влияют на процессы выделения нейросекрета.
Строение и функция промежуточной доли гипофиза.
Промежуточная часть (доля) аденогипофиза представлена эпителием, расположенным в несколько слоев, локализован¬ных между передней и задней долями гипофиза. В промежу¬точной части есть псевдофолликулы, содержащие коллоидоподобную массу. Функции: 1) секреция меланотропного (меланоцитостимулирующего) гормона, регулирующего обмен пигмента меланина; 2) липотропного гормона, регулирующе¬го обмен липидов.
Строение и функция нейрогипофиза. Источники развития.
нейрогипофиз (задняя доля).
Задняя доля гипофиза (нейрогипофиз) представлена в ос¬новном эпендимной глией. Клетки нейроглии называются питуицитами. В нейрогипофизе гормоны не вырабатывают¬ся (это нейрогемальный орган). В заднюю долю поступают аксоны нейросекреторных клеток супраоптического и паравентрикулярного ядер. По этим аксонам в заднюю долю транспортируются вазопрессин и окситоцин и накапливают¬ся на терминалях аксонов около кровеносных сосудов (является депо-резервуаром данных гормонов). Эти накопления называются накопительными тельцами, или тельцами Херринга. По мере надобности из этих телец гормо¬ны поступают в кровеносные сосуды.
ИСТОЧНИКИ РАЗВИТИЯ
• вырост из эктодермального эпителия первичной полости рта (карман Ратке) - передняя и промежуточная доли
• вырост из промежуточного мозгового пузыря (нервная трубка) - задняя доля
• мезенхима - капсула и прослойки соединительной ткани
Строение щитовидной железы. Источники развития.
Щитовидная железа - самая крупная из эндокринных желез организма - имеет у млекопитающих сложный гистогенез и вырабатывает тиреоидные гормоны (которые регулируют активность метаболических реакций и процессы развития), а также кальцитонин - гормон, участвующий в регуляции кальциевого обмена. Каждая из двух долей щитовидной железы покрыта капсулой из плотной волокнистой ткани, от которой внутрь органа отходят прослойки, несущие сосуда и нервы.
ЩИТОВИДНАЯ ЖЕЛЕЗА – эндокринная железа, имеет форму бабочки, расположена в передней части шеи и состоит из двух долей и перешейка, расположенного на уровне ΙΙ—ΙV хряща трахеи и соединяет между собой обе доли. Вес железы: 20 — 30 г. На задней поверхности двух долей парами расположены четыре околощитовидные железы. Снаружи щитовидная железа покрыта мышцами шеи, расположенными ниже подъязычной кости; своим фасциальным мешком железа прочно соединена с трахеей и гортанью, поэтому она перемещается вслед за движениями этих органов. Железа состоит из фолликулов — пузырьков овальной или округлой формы, которые заполнены белковым йодсодержащим веществом типа коллоида; между пузырьками располагается рыхлая соединительная ткань.
Эмбриональное развитие. Зачаток щитовидной железы возникает у зародыша человека на 3-4-й неделе как выпячивание стенки глотки между I-ой и II-ой парами жаберных карманов, которое растет вдоль глоточной кишки в виде эпителиального тяжа. На уровне III-IV пар жаберных карманов этот тяж раздваивается, давая начало формирующимся правой и левой долям щитовидной железы.
Строение и функция фолликула щитовидной железы.
Фолликулы - морфофункциональные единицы железы - замкнутые образования округлой формы, стенка которых состоит из одного слоя эпителиальных клеток (тироцитов), а в просвете содержится их секреторный продукт - коллоид (рис. 3-9). Каждый фолликул окружен капиллярной сетью в виде корзиночки. Размеры фолликулов у взрослого варьируют в пределах 50500 мкм; их количество, по разным оценкам, составляет от 3 до 30 млн.
Говоря о функциях фолликул мы говорим о функциях тироцитов
Функция тироцитов заключается в синтезе и выделении йодсодержащих тиреоидных гормонов - три- (Т3) и тетрайодтиронина (Т4) или тироксина.
Тироксин (Т4) и трийодтиронин (Т3) регулируют обмен веществ, рост и развитие организма. Синтез этих гормонов происходит в эпителиальных фолликулярных клетках, называемых тироцитами
Т4 и Т3 участвуют в регуляции метаболических реакций, влияют на рост и дифференцировку тканей, особенно на развитие нервной системы. Все этапы деятельности тироцитов находятся под контролем ТТГ, эффекты которого опосредуются рецепторами ТТГ на их плазмолемме.
Характеристика тироцитов.
Тироциты изменяют свою форму от плоской до цилиндрической в зависимости от функционального состояния. В норме у человека преобладают кубические клетки. Форма ядер соответствует форме клетки. Цитоплазма содержит хорошо развитые синтетический и лизосомальный аппараты, на апикальной поверхности имеются многочисленные микроворсинки, латеральная образует комплексы соединений, препятствующих утечке коллоида (на который могут вырабатываться антитела) из фолликулов.
Функция тироцитов заключается в синтезе и выделении йод-содержащих тиреоидных гормонов - Т3, или трийодтиронина, и Т4, или тироксина.
-В тироцитах хорошо развиты органеллы, особенно участвующие в белковом синтезе. Белковые продукты, синтезируемые тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиронинов (т.е. аминокислот, входящих в состав крупной и сложной молекулы тироглобулина). Тироидные гормоны могут попасть в циркуляцию лишь после высвобождения из этой молекулы (т.е. после расщепления тироглобулина)
Характеристика С-клеток.
С-клетки вырабатывают гормон кальцитонин, оказывающий гипокальциемическое действие. Они выявляются специальными способами окраски и наиболее часто лежат поодиночке или мелкими группами парафолликулярно - в стенке фолликула между тироцитами и базальной мембраной (рис. 3-10). У человека в норме их содержание в органе невелико (0.1% от общего числа клеток). Кальцитонин накапливается в
С-клетхах в плотных uранулах и выводится из клеток механизмом экзоцитоза при повышении уровня Са2+ в крови.
Строение и функция околощитовидной железы.
Околощитовидные железы выработают полипептидный паратгормон (паратирин), который участвует в регуляции кальциевого обмена.
Каждая из имеющихся в норме четырех желез покрыта тонкой капсулой из плотной соединительной ткани, от которой отходят перегородки, разделяющие ее на дольки. Тонкие прослойки соединительной ткани внутри долек содержат сеть фенестрированных капилляров, а также жировые клетки, число которых существенно нарастает с возрастом (у пожилых они нередко занимают 60-70% объема органа).
Паренхима железы представлена тяжами и скоплениями эпителиальных клеток (паратироцитов); иногда встречаются мелкие фолликулы с оксифильным содержимым. Выделяют два ведущих типа паратироцитов - главные и оксифильные.
1. Главные паратироцты (образуют основную часть паренхимы органа) - мелкие, полигональные клетки со слабо оксифильной цитоплазмой.
Встречаются в двух вариантах (светлые и темные клетки), отражающих различные функциональные состояния. Темные паратироциты - активно функционирующие клетки, с более развитыми грЭПС и комплексом Гольджи. Светлые (неактивные) содержат больше гликогена, лизосом, липидных капель и секреторных гранул диаметром 150-200 нм. Соотношение между темными и светлыми клетками в норме у человека составляет 1: 3-5.
Главные клетки осуществляют биосинтез и выделение паратгормона; регуляция этих процессов опосредуется их ионочувствительными механизмами: они угнетаются при повышении уровней Са2+ в крови (с накоплением в цитоплазме секреторных гранул) я активируются при их снижении.
Паратгормон увеличивает содержание и функциональную активность остеокластов в костной ткани и стимулирует ре абсорбцию кальция в почечных канальцах.
2.Оксифильные паратироциты располагаются среди главных и крупнее их, а их гиперхромные ядра имеют меньшие размеры. Цитоплазма интенсивно окрашивается кислыми красителями и отличается очень высоким содержанием крупных митохондрий при слабом развитии других органелл и отсутствии секреторных гранул. У детей эти клетки единичны, с возрастом их число нарастает.
Источники развития и функциональная роль оксифильных клеток до конца не выяснены. Мнение о том, что они являются дегенеративно измененными главными и не обладают специфической функцией, оспаривается рядом авторов.
Строение и функции коркового вещества надпочечника. Источники развития
Развитие надпочечников проходит в несколько этапов.
Закладка корковой части появляется на 5-й неделе внутриутробного периода в виде утолщений целомического эпителия. Эти эпителиальные утолщения собираются в компактное интерреналовое тело, - зачаток первичной (фетальной) коры надпочечников. С 10-й недели внутриутробного периода клеточный состав первичной коры постепенно замещается и дает начало дефинитивной коре надпочечников, окончательное формирование которой происходит в течение первого года жизни.
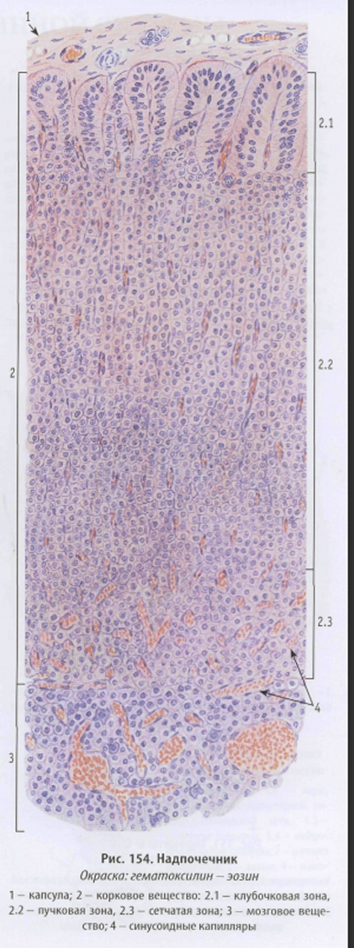
Корковое вещество образовано тремя нерезко разграниченными зонами:
(1)клубочковой - тонкой наружной, лежащей под капсулой; (2) пучковой - средней, образующей основную массу коры, и (3) сетчатой - узкой внутренней, прилежащей к мозговому веществу (рис. 3-12).
1. Клубочковая зона образована небольшими клетками с равномерно окрашенной цитоплазмой, которые формируют округлые арки ("клубочки"). В этих клетках хорошо развиты аЭПС и комплекс Гольджи; митохондрии - с пластинчатыми и тубулярными кристами. Липидных капель меньше, чем в пучковой зоне.
2. Пучковая зона состоит из крупных оксифильных вакуализированных клеток (спонгиоцитов), которые образуют радиально ориентированные тяжи ("пучки"), разделенные синусоидными капиллярами. Для них характерно высокое содержание липидных капель, митохондрии с тубулярными кристами, мощное развитие аЭПС и комплекса Гольджи (рис. 3-12).

Рис. 3-12. Надпочечник: вид на гистологическом препарате и ультраструктура клеток различных отделов. К - капсула, КЗ - клубочковая зона, ПЗ - пучковая зона, СЗ - сетчатая зона, МВ - мозговое вещество, КАП - капилляры.
3. Сетчатая зона образована анастомозирующими эпителиальными тяжами, идущими в различных направлениях, между которыми располагаются кровеносные капилляры. Клетки этой зоны меньших размереров, чем в пучковой зоне; в цитоплазме содержатся митохондрии с тубулярными кристами, аЭПС хорошо развита. Липидных капель меньше, чем в пучковой зоне, встречаются многочисленные лизосомы и липофусцыновые гранулы. Часты картины гибели клеток механизмом апоптоза.
Гормоны коркового вещества надпочечника - кортикостероиды -
разделяются на три основных класса, которые синтезируются и выделяются в клубочковой, пучковой и сетчатой зонах коркового вещества, соответственно:
1.Минералкортнкоиды - влияют на уровни электролитов в крови и артериальное давление (у человека наиболее важен из них альдостерон);
2.Глюкокортикоиды - оказывают выраженное действие на различные виды обмена (особенно углеводный) и на иммунную систему (главным из них у человека является кортизол);
3.Половые стероиды (главные из них у человека -
дегидроэпиандростерон и его сульфат - имеют слабое андрогенное действие).
Исходным субстратом для синтеза всех кортикостероидов служит
холестерин, который путем рецепторно-опосредованного эндоцитоза
извлекается клетками из крови в составе липопротеинов низкой плотности, отщепляется от них после гидролиза в лизосомах и накапливается в липидных каплях. Ферментные системы, участвующие в синтезе стероидных гормонов (стероидогенезе), расположены в аЭПС и митохондриях. Стероиды не запасаются в клетках, а образуются и выделяются непрерывно.
Регуляция деятельности клеток коры надпочечника в пучковой и сетчатой зонах обеспечивается АКТГ, взаимодействующим со специфическим рецептором на их плазмолемме. Секреция АКТГ. в свою очередь, контролируется кортиколиберином гипоталамуса. Синтез и секреция минералкортикоидов клетками клубочковой зоны регулируются преимущественно ренин-ангиотензиновой системой.
Строение и функции мозгового вещества надпочечника. Источники развития.
Мозговое вещество образовано хромаффинными, ганглиозными и поддерживающими клетками.
Мозговое вещество надпочечников вырабатывает катехоламины (адреналин и норадреналин), которые поддерживает тонус симпатической нервной системы, оказывают выраженное стимулирующее влияние на мышечную работоспособность; стимулируют синтез стероидных гормонов. Адреналин часто называют гормоном борьбы или бегства, так как его выделение резко возрастает лишь в минуты опасности. Повышение уровня адреналина в крови влечёт за собой соответствующие физиологические изменения — учащается сердцебиение, сужаются кровеносные сосуды, напрягаются мышцы, расширяются зрачки. Ещё корковое вещество в небольших количествах вырабатывает мужские половые гормоны (андрогены).
а) хромаффинные клетки - основной элемент мозгового вещества - расположены в виде гнезд и тяжей, имеют полигональную форму, крупное ядро, мелкозернистую или вакуолизированную цитоплазму. Гистохимическими методами выделяют два их типа: Н- и (численно
преобладающие) А-клетки, вырабатывающие норадреналин и адреналин, соответственно.
Содержат мелкие митохондрии, ряды цистерн грЭПС, крупный комплекс Гольджи, многочисленные секреторные гранулы - с умеренно плотным (в А-
клетках) или уплотненным в центре и светлым по периферии (в Н-клетках)
матриксом. Секреторные гранулы помимо катехоламинов содержат белки (в том числе хромогранины - осмотические стабилизаторы - и энкефалины, ИЛ-1, факторы роста), липиды и АТФ
б) ганглиозные клетки содержатся в небольшом числе и представляют собой вегетативные нейроны;
в) поддерживающие клетки – отростчатые, глиальной природы: охватывают хромаффинные клетки.
Физиологическая регенерация коркового вещества надпочечника описывается двумя теориями: миграционной и зональной.
1.Миграционная гипотеза более обоснована, базируется на представлении
отом, что новообразование клеток коркового вещества происходит в клубочковой зоне, откуда они мигрируют в пучковую и далее - в сетчатую, где подвергаются дегенеративным изменениям и гибнут. По ходу миграции клетки претерпевают фенотипические изменения и продуцируют различные классы кортикостероидов.
2.Зональная гипотеза предполагает сравнительную независимость каждой из зон, обеспечивающих необходимый уровень своей регенерации.
Понятие о диффузной эндокринной системе.
Диффузная эндокринная система (ДЭС) образована эндокриноцитами, рассеянными по различным органам и выявляемыми при использовании специальных методов окраска (в частности, солей серебра). Клетки ДЭС располагаются поодиночке или мелкими группами. Значительное их число находится в слизистых оболочках различных органов и связанных с ними железах. Они особенно многочисленны в пищеварительном тракте (гастро- энтеро-панкреатическая система).
Клетки ДЭС в слизистых оболочках (рис. 3-13) имеют широкое основание и более узкую апикальную часть, которая в одних случаях доходит до просвета органа (клетки открытого типа), а в других с ним не контактирует (клетки закрытого типа). Предполагается, что эти клети участвуют в анализе химического состава пищи, воздуха, мочи и т.п. и отвечают на его изменения выделением гормонов и паракринных факторов.
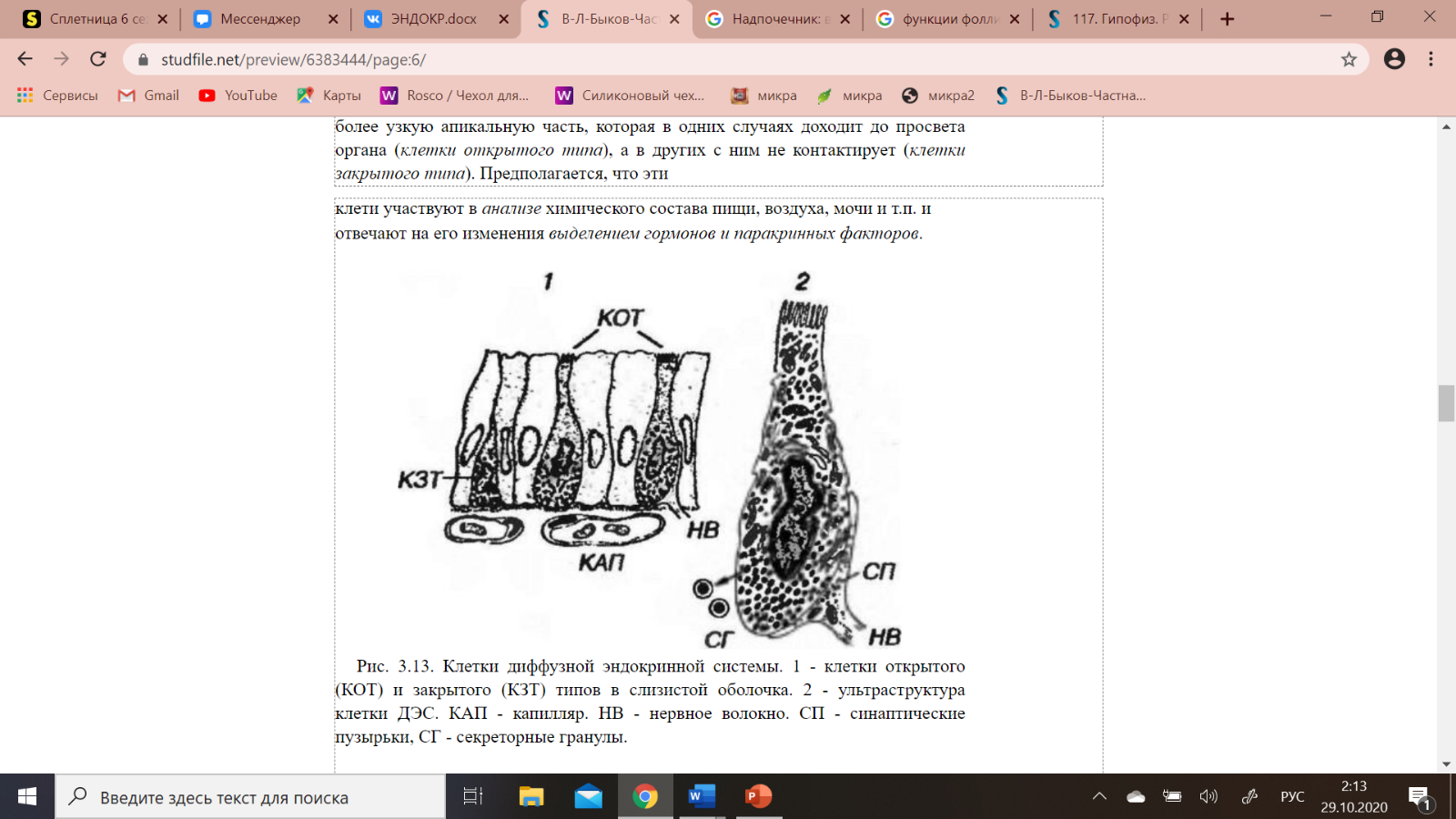
Рис. 3.13. Клетки диффузной эндокринной системы. 1 - клетки открытого (КОТ) и закрытого (КЗТ) типов в слизистой оболочка. 2 - ультраструктура клетки ДЭС. КАП - капилляр. НВ - нервное волокно. СП - синаптические пузырьки, СГ - секреторные гранулы.
Клетки ДЭС характеризуются сравнительно слабым развитием грЭПС и комплекса Гольджи; для них типично наличие аргирофильных плотных секреторных гранул в базальных отделах цитоплазмы.
Секреторные продукты клеток ДЭС оказывают как местные
(паракринные), так и дистантные (эндокринные) влияния. Они синтезируют и выделяют ряд структурно родственных пептидов и биоаминов, которые играют роль нейромедиаторов и гормонов. Эффекты этих веществ очень многообразны, в частности, они влияют на моторику гладкомышечной ткани в стенке разных органов или секрецию экзо- и эндокринных желез (см., например, 95).
Представление о ДЭС тесно смыкается с понятием APUD-системы, названной так по первым буквам английских терминов, характеризующих функциональные свойства клеток этой системы - Amine Precursor Uptake and Decarboxylation - способность к захвату предшественников аминов и (их) декарбоксилированию.
ОРГАНЫ КРОВЕТВОРЕНИЯ И ИММУНОГЕНЕЗА
ПРЕПАРАТЫ
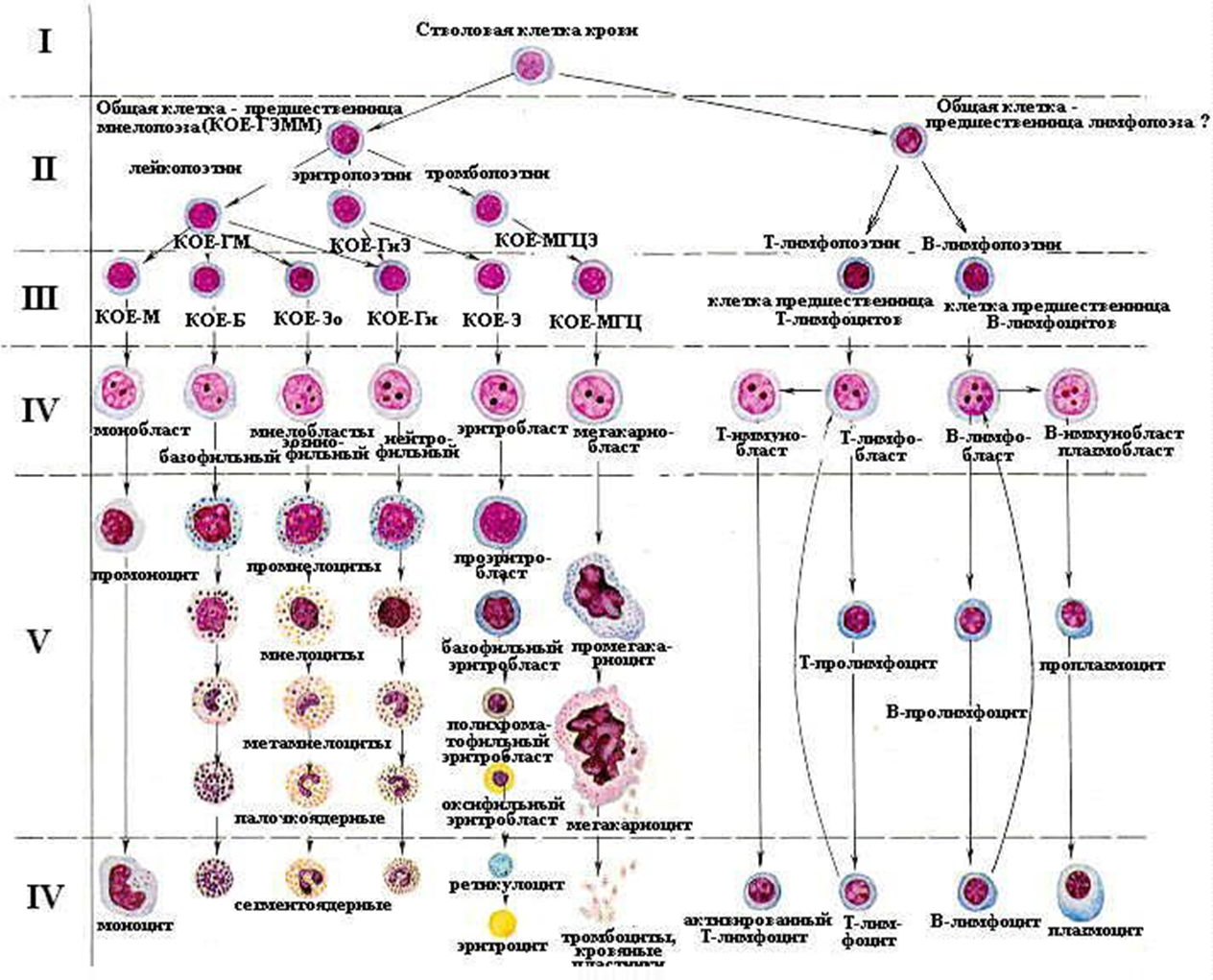
Гемопоэз — процесс образования форменных элементов крови: эритроцитов (эритропоэз), лейкоцитов (лейкопоэз) и тромбоцитов (тромбоцитопоэз).
Он совершается в красном костном мозге, где образуются эритроциты, все зернистые лейкоциты, моноциты, тромбоциты, В-лимфоциты и предшественники Т-лимфоцитов. В тимусе проходит дифференцировка Т-лимфоцитов, в селезенке и лимфатических узлах — дифференцировка В-лимфоцитов и размножение Т-лимфоцитов.
1 класс — стволовая полипотентная клетка, способная к поддержанию своей популяции. По морфологии соответствует малому лимфоциту, является полипотентной, то есть способной дифференцироваться в любой форменный элемент крови. Направление дифференцировки стволовой клетки определяется уровнем содержания в крови данного форменного элемента, а также влиянием микроокружения стволовых клеток — индуктивным влиянием стромальных клеток костного мозга или другого кроветворного органа. Поддержание численности популяции стволовых клеток обеспечивается тем, что после митоза стволовой клетки одна из дочерних клеток становится на путь дифференцировки, а другая принимает морфологию малого лимфоцита и является стволовой. Делятся стволовые клетки редко (1 раз в полгода), 80 % стволовых клеток находятся в состоянии покоя и только 20 % в митозе и последующей дифференцировке. В процессе пролиферации каждая стволовая клетка образует группу или клон клеток и потому стволовые клетки в литературе нередко называются колоние—образующие единицы — КОЕ.
2 класс — полустволовые, ограниченно полипотентные (или частично коммитированные) клетки—предшественницы миелопоэза и лимфопоэза. Имеют морфологию малого лимфоцита. Каждая из них дает клон клеток, но только миелоидных или лимфоидных. Делятся они чаще (через 3—4 недели) и также поддерживают численность своей популяции.
3 класс — унипотентные поэтин—чувствительные клетки—предшественницы своего ряда кроветворения. Морфология их также соответствует малому лимфоциту. Способны дифференцироваться только в один тип форменного элемента. Делятся часто, но потомки этих клеток одни вступают на путь дифференцировки, а другие сохраняют численность популяции данного класса. Частота деления этих клеток и способность дифференцироваться дальше зависит от содержания в крови особых биологически активных веществ — поэтинов, специфичных для каждого ряда кроветворения (эритропоэтины, тромбопоэтины и другие).
Первые три класса клеток объединяются в класс морфологически неидентифицируемых клеток, так как все они имеют морфологию малого лимфоцита, но потенции их к развитию различны.
4 класс — бластные (молодые) клетки или бласты (эритробласты, лимфобласты и так далее). Отличаются по морфологии как от трех предшествующих, так и последующих классов клеток. Эти клетки крупные, имеют крупное рыхлое (эухроматин) ядро с 2 4 ядрышками, цитоплазма базофильна за счет большого числа свободных рибосом. Часто делятся, но дочерние клетки все вступают на путь дальнейшей дифференцировки. По цитохимическим свойствам можно идентифицировать бласты разных рядов кроветворения.
5 класс — класс созревающих клеток, характерных для своего ряда кроветворения. В этом классе может быть несколько разновидностей переходных клеток — от одной (пролимфоцит, промоноцит), до пяти в эритроцитарном ряду. Некоторые созревающие клетки в небольшом количестве могут попадать в периферическую кровь (например, ретикулоциты, юные и палочкоядерные гранулоциты).
+6 класс — зрелые форменные элементы крови. Однако следует отметить, что только эритроциты, тромбоциты и сегментоядерные гранулоциты являются зрелыми конечными дифференцированными клетками или их фрагментами. Моноцитыне окончательно дифференцированные клетки. Покидая кровеносное русло, они дифференцируются в конечные клетки — макрофаги. Лимфоциты при встрече с антигенами, превращаются в бласты и снова делятся.

КРАСНЫЙ КОСТНЫЙ МОЗГ


Уметь определять и объяснять строение: 1 – миелоидная ткань; 1.1 – ретикулярные клетки; 1.2 – липоциты; 1.3 – клетки-бласты; 1.4 – миелоцит; 1.5 нормоцит; 1.6 – мегакариоцит; 1.7 – зрелые форменные элементы крови; 2 – синусоидные капилляры.
1 - кость:1.1 - компактное вещество, 1.1.1 - сосуд, проникающий в губчатое вещество, 1.2 - губчатое вещество, 1.2.1 - костные трабекулы, 1.2.2 - эндост;2 - гемопоэтический компонент;
3 - стромальный компонент: 3.1 - ретикулярные клетки, 3.2 - жировые клетки, 3.3 - макрофаги с гранулами кармина в цитоплазме; 4 - сосудистый компонент: 4.1 - синусоиды (венулярные синусы), 4.2 - центральная вена
 аа
аа
ТИМУС
Уметь определять и объяснять строение: 1 – долька; 1.1 – корковое вещество; 1.2 – мозговое вещество; 1.2.1 – лимфоциты; 1.2.2 – эпителио-ретикулярные клетки; 1.2.3 – тимусное тельце; 2 – междольковая соединительная ткань; 3 – капсула.



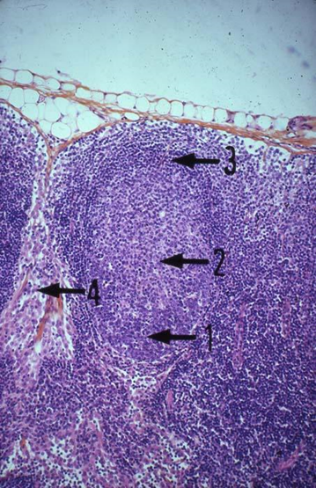
ЛИМФАТИЧЕСКИЙ УЗЕЛ
Уметь определять и объяснять строение: капсула; рабекула; лимфатические синусы: краевой; корковый; мозговой; корковое вещество; лимфоидный узелок; герминативный центр; венец; паракортикальная зона; мозговое вещество; мозговые тяжи.









СЕЛЕЗЕНКА


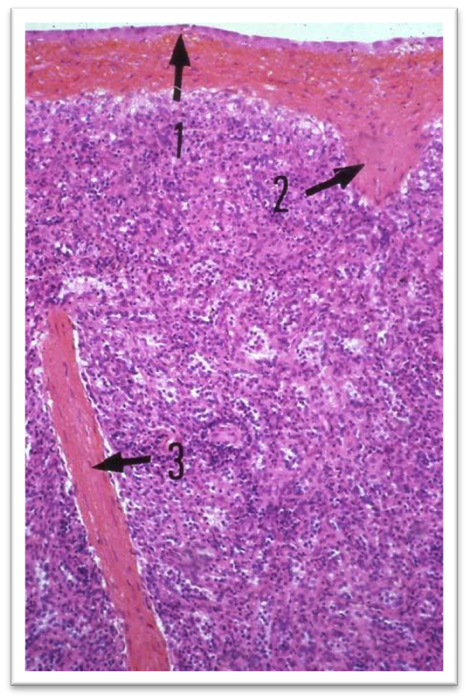

Уметь определять и объяснять строение: капсула; гладкомышечные клетки; мезотелий; трабекула; трабекулярная артерия; трабекулярная вена; белая пульпа; центральная артерия; лимфоидная муфта; лимфоидный узелок; маргинальная зона

ТЕРМИНЫ
Антиген-независимая пролиферация - В процессе дифференцировки Т-лимфоцитов выделяют два основных этапа (как Вы помните, такие же два этапа выделяют в процессе дифференцировки В-лимфоцитов): Антигеннезависимая дифференцировка – происходит постоянно в тимусе.
Антиген-зависимая пролиферация - Антигензависимая пролиферация и дифференцировка Т- и В-лимфоцитов происходят при встрече с антигенами в периферических лимфоидных органах, при этом образуются эффекторные клетки и клетки памяти (сохраняющие информацию о действовавшем антигене). происходит в периферических органах иммунной системы только при контакте Т-лимфоцита с антигеном.
Акцидентальная инволюция (ТИМУСА) - представляет собой быстрое уменьшение массы железы, под влиянием глюкокортикостероидов при различных стрессовых состояниях, при многочисленных инфекционных заболеваниях, лейкозах, злокачественных опухолях.
Береговые клетки - ретикулоэндотелиальными ("береговыми") клетками, между которыми во многих местах находятся оседлые макрофаги.
Белая пульпа - лимфоидная ткань, включающая 2 компонента: периартериальные влагалища - скопления Т-лимфоцитов вокруг пульпарных артерий, и лимфатические узелки, или фолликулы (1 на снимках в-г), содержащие и В-, и Т-клетки.
В-зависимая зона - это зоны лимфоидных фолликулов лимфоузлов, селезенки, аппендикса, миндалин, лимфоидных агрегатов кишечника.
Вторичный лимфатический узелок - через несколько лет после рождения все лимфатические узелки лимфоузлов становятся вторичными: это значит, что они
когда-то подверглись антигенной стимуляции.
Гемопоэтический компонент красного костного мозга - образован миелоидной тканью и содержит клетки миелоцшпарного и лимфоцитарного рядов на разных стадиях развития, взаимодействующие со стромальными эле¬ментами. В нем находится самоподдерживающаяся популяция плюрипо-тентных стволовых клеток (1/2000 клеток мозга).
Гемато-тимусный барьер - Гема́то-ти́мусный барьер — физиологический гистогематический барьер между кровеносной системой и тимусом, обеспечивающий защиту созревающих T-лимфоцитов от контакта с антигенами.
Герминативный центр - зона лимфоидного узелка (фолликула) периферической лимфоидной ткани, в которой зрелые В-лимфоциты, активированные антигеном, пролиферируют, дифференцируются и претерпевают процессы соматического гипермутагенеза и переключения классов антител.
Гуморальные иммунные реакции - иммунная реакция, осуществляемая посредством В-лимфоцитов и включающая производство антител. Гуморальный иммунитет связан с выработкой цитокинов интерлейкина-4 и интерлейкина-10.
Иммунокомпетентные клетки - это клетки, обеспечивающие функционирование иммунной системы. К иммунокомпетентным клеткам относятся: Антигенпрезентирующие клетки (АПК); Т-лимфоциты; В-лимфоциты; NK-клетки (клетки-киллеры).
Иммуноглобулины - это группа белков плазмы крови, синтезируемых В- лимфоцитами и плазмоцитами.
Интерлейкин(ы) - группа цитокинов, синтезируемая в основном лейкоцитами (по этой причине был выбран корень «лейкин»). Также производятся мононуклеарными фагоцитами и другими тканевыми клетками. Интерлейкины являются частью иммунной системы.
Клеточные иммунные реакции - такой тип иммунного ответа, в котором не участвуют ни антитела, ни система комплемента. В процессе клеточного иммунитета активируются макрофаги, натуральные киллеры, антиген-специфичные цитотоксические Т-лимфоциты, и в ответ на антиген выделяются цитокины.
Колониестимулирующий фактор –- один из основных препаратов, используемых в онкологической практике для лечения нейтропении.
«Клетки-няньки»коркового вещества тимуса - имеют глубокие инвагинации, в которых и происходит развитие Т-клеток.
Корона лимфатического узелка - скопление малых Влимфоцитов, расположенных по периферии фолликула, содержит В клетки памяти, незрелые плазматические клетки. Герментативный центр - в нем происходит пролиферация и дифференцировка В-клеток в незрелые плазматические и В-клетки памяти в результате их взаимодействия с антигеном.
Красная пульпа - содержимое пространства между лимфоидной тканью и трабекулами. плазмоциты. тоже заполненные клетками крови.
Лимфатический узелок - это скопление В-лимфоцитов, имеющее округлую форму. Функционально лимфатические узелки - это В-зона. Функция - в них происходит антигенЗАВИСИМАЯ пролиферация и дифференцировка В-лимфоцитов. Ее результат - «наивные» В-лимфоциты дифференцируются в плазмоциты. Плазмоциты (плазматические клетки) секретируют все 5 классов иммуноглобулинов (антител).
Лимфатические синусы - пространства, выстланные ретикулоэндотелиальными ("береговыми") клетками и служащие для перемещения лимфы через узел.
Межузелковая зона - содержит малые лифоциты и макрофаги, в условиях антигенной стимуляции она исчезает.
Мозговой синус - между трабекулами и мозговыми тяжами. (????????)
Отрицательная селекция тимоцитов - В тимусе производятся белки со всего организма (включая, к примеру, инсулин), правда, в мизерных количествах — только для того, чтобы представить их на поверхности APC. Если Т-клетка распознает такой белок — она сразу же уничтожается; ошибка в процессе уничтожения обычно приводит к аутоиммунным заболеваниям (в случае с инсулином — к диабету первого типа).
Периферические органы кроветворения и иммуногенеза - селезенка; лимфатические узлы (у млекопитающих и водоплавающих птиц), железа Гардера, слезная железа, дивертикул Меккеля (у птиц); диффузная и узелковая лимфоидная ткань органов пищеварения, дыхания, кожи, мочеполовых путей, сальника, миндалины. Это центральный орган кроветворения и иммунной системы.
Плюрипотентные стволовые клетки -Плюрипотентные клетки могут дифференцироваться во все типы клеток, кроме клеток внезародышевых органов (плаценты и желточного мешка).
Посткапиллярные синусы – функциональные сосуды красного костного мозга.
Претимоциты – предшественники Т-лимфоцитов
Положительная селекция тимоцитов - В процессе развития тимоцитов происходит увеличение популяции клеток, экспрессирующих Т-клеточные рецепторы, распознающие чужеродные антигены (положительная селекция), а также удаление популяций тимоцитов, реагирующих с аутодетерминантами (отрицательная селекция).
Периваскулярные клетки коркового вещества тимуса - охватывают уплощенными отростками капилляры и служат элементом гемато-тимусного барьера (имеется только в корковом веществе), предотвращающего воздействие циркулируюших в крови антигенов на созревающие зямоциты.
Первичный лимфатический узелок - компактные однородные скопления малых В-лимфоцитов, связанных с ретикулярными клетками. Имеется небольшое число Т лимфоцитов, макрофагов. Под влиянием антигенов превращаются во вторичные. Вторичные узелки состоят из короны и герментативного центра.
Паракортикальная зона - Глубокая кора (паракортикальная зона) является Т-зависимой зоной, в которой осуществляется дозревание т клеток, поступивших из тимуса, а также их антиген зависимая пролиферация и дифференцировка.
Промежуточный синус - корковые (промежуточные) синусы, которые окружают лимфоидные узелки
Периартериальное лимфатическое влагалище - скопления Т-лимфоцитов вокруг пульпарных артерий, и лимфатические узелки, или фолликулы, содержащие и В-, и Т-клетки.
Стромальный компонент красного костного мозга - строма, служащая микроокружением для гемопоэтических (кроветворных) клеток, и гемального — форменные элементы крови на разных стадиях развития.
Сосудистый компонент красного костного мозга - представлен многочисленными кровеносными сосудами. Наряду с обычными компонентами микроциркуляторного русла содержит особые посткапиллярные синусы. Артерии, проникая в костномозговую полость, распадаются на многочисленные капилляры, которые переходят в венозные синусы.
Субкапсулярная зона - Здесь сосредоточено 5% Т-лимфоцитов, которые устойчивы (резистентны) к физическим факторам, облучениям и глюкокортикоидам коры надпочечников. При облучении или при стрессе лимфоциты данной зоны меньше всего страдают, в то время как остальные лимфоциты могут разрушаться (при стрессе).
Секреторные клетки коркового вещества тимуса - выделяют факторы, стимулирующие Т-лимфоцитопоэз.
Субкапсулярный синус - После попадания в лимфатический узел из афферентных лимфатических сосудов лимфа течет в пространство под капсулой, называемое субкапсулярным синусом
Т-зависимая зона - это паракортикальная зона лимфоузлов, периартериальные зоны селезенки, интерфолликулярные зоны в аппендиксе, миндалинах, пейеровых бляшках.
Тимоцит - общее название клеток вилочковой железы.Эпителиальная клетка.
Тельца Гассаля - (тельца вилочковой железы, "эпителиальные жемчужины") — слоистые эпителиальные тельца, расположенные в средней части мозгового вещества долек тимуса, образованы концентрически наслоёнными, плотными, сильно уплощёнными эпителиальными клетками.
Тяжи Бильрота - Пульпарные тяжи.
Часть красной пульпы, расположенная между синусами, называется селезеночными, или пульпарными, тяжами (chordae splenicae) Бильрота. Это форменные элементы крови, макрофаги, плазматические клетки лежащие в петлях ретикулярной соединительной ткани.
Эритропоэтин - один из гормонов почек (также секретируется в перисинусоидальных клетках печени), который контролирует эритропоэз, то есть образование красных кровяных клеток (эритроцитов). По химическому строению является гликопротеином.
Эндост - тонкий соединительнотканный слой, выстилающий костную ткань трубчатых костей изнутри с формированием костномозгового канала. Морфологически эндост подобен наружному слою трубчатых костей — периосту (надкостнице).
ВОПРОСЫ
1.Классификация органов кроветворения и иммунных реакций.
Иммунные реакции разделяются на:
1)клеточные, обусловленные непосредственным контактным воздействием иммунокомпетентных клеток на их мишени (осуществляются Т-лимфоцитами, взаимодействующими с аитиген-представляющими клетками);
2)гуморальные, связанные с выработкой циркулирующих в крови и жидкостях организма антител - иммуноглобулинов, которые нейтрализуют антигены (обеспечиваются В-лимфоцитами, взаимодействующими с антиген представляющими клетками и Т-лимфоцитами).
Центральные органы (красный костный мозг, тимус) обеспечивают процессы антиген-независимой пролиферации и дифференцировки клеток-предшественников, поступающих из красного костного мозга. При этом образуются клетки с огромным репертуаром рецепторов ко всевозможным антигенам. Такое разнообразие обусловлено реаранжировкой их генома; антигены на этом этапе не только не нужны, но даже вредны.
Периферические органы (все остальные органы иммунной системы) обеспечивают процессы антиген-зависимой пролиферации и дифференцировки клеток, мигрирующих из центральных органов, где они ранее приобрели специфические рецепторы к данному антигену. Для обеспечения контакта с антигенами эти органы расположены на путях их поступления через лимфу или кровь.
2.Строение красного костного мозга. Источники развития.
Красный костный мозг представляет собой центральный орган кроветворения и иммуногенеза, содержащий самоподдерживающуюся популяцию стволовых клеток крови и участвующий в образе лини клеток миелоцитарного и лимфоцитарного рядов. У взрослого он рассредоточен по всему организму (что маскирует его массу - 1.5-2 кг), находится в ячейках губчатого вещества костей (в плоских костях и эпифизах трубчатых костей).
В состав красного костного мозга входят три компонента: 1)гемопоэтический, 2) стромальный, 3) сосудистый

1. Гемопоэтический компонент образован миелоидной тканью и содержит клетки миелоцитарного и лимфоцитарного рядов на разных стадиях развития, взаимодействующие со стромальными элементами. В нем находится самоподдерживающаяся популяция плюрипотентных стволовых клеток (1/2000 клеток мозга).
2.Стромальный компонент (функции см. выше) включает: ретикулярные клетки отростчатой формы и волокна, образующие трехмерную сеть (ретикулярные клетки, прилежащие к стейке синусов, называют адвентициальными); адипоциты (жировые клетки); макрофаги; клетки эндоста (соединительнотканной выстилки костных полостей);
3.Сосудистый компонент наряду с обычными сосудами микроциркуляторного русла содержит особые посткапилпярные (венозные) синусы - тонкостенные анастомозирующие сосуды диаметром 50-75 мкм. Синусы выстланы тонким эндотелием, способным отличать зрелые форменные элементы гемопоэтического компонента от незрелых и пропускать их в просвет синуса через временно образующиеся в цитоплазме клеток поры. Базальная мембрана на большем протяжении отсутствует. Наружный (прерывистый) слой стенки синусов образуют адвентициальные клетки. Синусы снабжены сфинктерами и способны временно выключаться из кровотока, играя роль "отстойников", в которых дозревают форменные элементы. К ним снаружи прилегают макрофаги, проникающие своими отростками в просвет синусов.
Распределение клеток различных типов в гемопоэтическом компоненте не случайно. Так, мегакариоциты всегда лежат вблизи синусов, проникая в их просвет своими отростками в виде лент, которые распадаются на отдельные тромбопиты. Гранулоциты созревают вблизи клеток эндоста и контактируют с ретикулярными клетками и преадипоцитами. Эритроидные элементы развиваются в составе т.н. эритробластических островков, в контакте с ретикулярными клетками, которые накапливают и передают им частицы ферритнна (содержащие железо, необходимое для синтеза гемоглобина).
Источники развития:
1) мезенхима – соед.тк., ретик. тк., сосуды
2) внезарод. мезенхима - СКК (мигр. из желт. мешка) – форменные элементы крови на разных стадиях развития.
3) нейроэктодерма. Закладывается на 2-м мес. внутр. развития. Как основной орган кроветворения начинает функционировать у 20-28-нед. плода
3.Функции красного костного мозга.
1) миелопоэз (миелоидное кроветворение) – эритроциты, гранулоциты (э.,б.,н.), моноциты, тромбоциты
2) защитная
3) аналог сумки Фабрициуса птиц – антигеннезависимая диффер. В-л/ц
4) образование унипот. клеток – предш. Т-л/ц
5) образование NK-клеток (натуральных, естественных киллеров)
Красный костный выполняет две главные функции:
· образование и дифференцировка всех клеток крови на основе самоподдерживающейся популяции стволовой клетки
· антигеннезависимая дифференцировка В-лимфоцитов. Источник развития - стволовая клетка.
Красный костный мозг помимо функции кроветворения выполняет также функцию имунногенеза, являясь центальным органом имуногенеза. В красном костном мозге происходит антиген-независимая пролиферация В-лимфоцитов. В ходе этого процесса В-лимфоциты на своей поверхности приобретают имунноглобулиновые рецепторы к разным антигенам. И в таком состоянии отправляются в антиген-зависимые зоны периферических органов кроветворения.
4.Строение тимуса. Источники развития.
Источники развития:
1) мезенхима – с/тканная строма, сосуды,
2) эпителий глоточной кишки в области 3-4 пар жаберных карманов – эпителиоретикулоциты
3) УПК предшественники Т-л – Т-л;
4) нейроэктодерма. Строение – дольчатое, корковое и мозговое в-во. В корковом – подкапсулярная зона. Строма С/тканная капсула, перегородки с сосудами, дольки. Основа стромы – ретикулоэпителиоциты – отростчатые, оксифильные клетки. Отростками охватывают лимфоцитарные клетки. Создают для них микроокружение, необходимое для их деления и созревания. Отростками клетки связаны друг с другом, создавая подобие синцития.
Тимус (зобная, или вилочковая, железа) представляет собой центральный орган иммунной системы, в котором происходит антиген-независимая пролиферация и дифференцировка Т-лимфоцитов из их предшественников, поступающих из красного костного мозга. Наибольшего развития достигает в детстве, после полового созревания подвергается возрастной инволюции, частично замещаясь жировой тканью.
Покрыт соединительнотканной капсулой, которая продолжается в перегородки, содержащие сосуды и разделяющие его на связанные друг с другом дольки. Долька состоит из трехмерной сети отростчатых эпителиальных (эпителиоретикулярных) клеток, образующих строму органа, в петлях которой располагаются лимфоциты (тимоциты). В каждой дольке выделяют корковое и мозговое вещество

5.Строение и функции коркового вещества дольки тимуса.
1. Корковое вещество - более темное вследствие плотной упаковки тимоцитов (содержит около 90% их числа). Предшественники Т-клеток (претимоциты) поступают в него из красного костного мозга, мигрируя через стенку сосудов кортико-медуллярной зоны; пролиферирующие тимоциты располагаются в виде скоплений между эпителиальными клетками в т.н. субкапсулярной зоне, имеют вид больших лимфоцитов и еще не обладают рецепторами Т-клеток (РТК). Созревающие тимоциты, продолжая делиться и перемещаясь в более глубокие части коры, за счет реаранжировки генома образуют РТК к различным антигенам. Они имеют вид средних и малых лимфоцитов. Тимоциты коры при стрессе разрушаются (под действием кортикостероидов), что вызывает опустошение коры (акцидентальную инволюцию).

Подавляющее большинство (90-95%) тимоцитов, образовавшихся в коре, в ней же гибнет механизмом апоптоза в процессе отбора, включающего положительную селекцию (выживание клеток, способных распознавать собственные белки главного комплекса гистосовместимости) и отрицательную селекцию (гибель клеток с рецепторами к собственным антигенам). Погибшие клетки уничтожаются макрофагами. В коре образуются субпопуляции Т-клеток: сначала они одновременно экспрессируют маркеры хелперов/индукторов (CD4), киллеров/супрессоров (CD8) и РТК, затем - РТК и только один из маркеров. Более зрелые Т-клетки перемешаются в мозговое вещество.
Эпителиоретикулярные клетки - светлые, оксифильные, со светлым ядром, крупным ядрышком и умеренно развитыми органеллами. Своими отростками они охватывают тимоциты, создавая микроокружение, необходимое для их деления и созревания (рис. 2-3). В корковом веществе имеется несколько особых вариантов эпителиальных клеток:
1)секреторные клетки (нескольких типов) - содержат в цитоплазме секреторные гранулы. Вырабатывают факторы, необходимые для созревания тимоцитов: тимозин, тимопоэтин, тимусный сывороточный фактор и др.
Первые два поступают также в кровь и способны оказывать действие вне тимуса, что позволяет рассматривать его как эндокринную железу;
2)"клетки-нянькии" - заключают в своей цитоплазме до нескольких десятков активно делящихся и часто гибнущих тимоцитов, изолируя их от окружающих клеток и участвуя, по-видимому, в их селекции;
3)периваскулярные клетки - охватывают уплощенными отростками капилляры и служат элементом гемато-тимусного барьера (имеется только в корковом веществе), предотвращающего воздействие циркулируюших в крови антигенов на созревающие зямоциты. В состав барьера входят (см. рис. 2-3):
(1)эндотелий капилляра (в некоторых участках также перициты);
(2)базальная мембрана капилляра;
(3)перикапилллрное пространство, содержащее волокна и макрофаги;
(4)базальная мембрана зпителиоретикулярных клеток;
(5)цитоплазма зпителиоретикулярных клеток, связанных десмосомами.
6.Гемато-тимусный барьер.
физиологический гистогематический барьер между кровеносной системой и тимусом, обеспечивающий защиту созревающих T-лимфоцитов от контакта с антигенами. Образован эпителием подкапсульных отделов тимуса и капиллярной сетью.
Особенности кровоснабжения тимуса. Гемато-тимусный барьер
Входящая в тимус вилочковая артерия делится на междольковые, а затем на внутридольквые артерии. На границе коркового и мозгового вещества последняя формирует дуговую артерию. Дуговая артерия образует 2 сети кровеносных капилляров. Наружная сеть капилляров уходит в корковое вещество, проходя между опорными клетками, эти капилляры участвуют в образовании гематотимусного барьера.
Он состоит:
1. Стенка капилляра: эндотелиоциты с непрерывной утолщенной базальной мембраной
2. Перикапиллярное пространство, представленное тканевой жидкостью, макрофагами и лимфоцитами.
3. Опорные эпителиоретикулоциты, или точнее их цитоплазма, и базальная мембрана этих клеток.
Этот барьер существует только в корковом веществе дольки, он предотвращает взаимодействие циркулирующих в крови антигенов и созревающих Т-лимфоцитов.
Глубокая сеть капилляров, представленная меньшим их количеством, уходит в мозговое вещество, не образуя барьера.
Отток осуществляется из коркового вещества в подкапсулярные вены, а из мозгового вещества – в посткапиллярные венулы с высоким эндотелием, а затем в междольковые вены.
7.Строение и функция мозгового вещества дольки тимуса.
Мозговое вещество - светлее коркового, содержит меньшее количество более зрелых (малых) тимоцитов, нечувствительных к кортикостероидам, которые покидают тимус (проходя через стенку посткапиллярной венулы в кортико-медуллярной зоне) и заселяют Т-зависимые зоны периферических органов иммунной системы. Эпителиальные клетки - более крупные и многочисленные, чем в коре; в отдельных участках они, уплощаясь и ороговевая, накладываются друг на друга концентрическими слоями, образуя слоистые эпителиальные тельца (Гассаля) диаметром до 100 мкм и более. Функция слоистых телец неясна; их размеры и число увеличиваются с возрастом и при стрессе.
мозговое вещество: образовано Т-лимфоцитами, макрофагами, иногда встречаются плазматические клетки
функции: точные функции неизвестны, возможно, какие-нибудь этапы антигеннезависимой дифференцировки Т-лимфоцитов
8.Строение и значение тимусных телец.
Особенностью мозгового вещества является наличие в центральной части телец Гассаля, которые образованны концентрически расположенными веретеновидными эпителиальными клетками на разных этапах зернистого перерождения, в центре тельца клетки содержат вещество близкое к кератину эпидермиса кожи. Слоистые тельца связаны с кровеносными сосудами. Они появляются на 12 неделе эмбриогенеза, наибольшее количество их в тимусе наблюдается в возрасте 15 лет, у пожилых людей число телец снижается, а в старческом наблюдаются крупные одиночные тельца диаметром 20-50 мкм. Назначение клеток в тельцах Гассаля не ясно, возможно, они выполняют эндокринную функцию или с их участием происходит гибель лимфоцитов.
Функция слоистых телец неясна; их размеры и число увеличиваются с возрастом и при стрессе.
9.Понятие о Т- и В- зависимых зонах.
Для периферических органов иммунной системы помимо этого характерны: В- и Т-зависимые зоны - участки с преимущественным расположением пролиферирующих и дифференцирующихся В- или Т- лимфоцитов, взаимодействующих со специфическими типами антигенпредставляющих клеток. В-зависимые зоны часто имеют вид лимфатических узелков - компактных округлых лимфоидных образований. С ними связаны диффузные скопления лимфоидной ткани; межфолликулярные участки лимфоидной ткали обычно соответствуют Т-зависимым зонам.
Наружная кора включает лимфоидную ткань, образующую лимфатические узелки (В-зависимые зоны) и межузелковые скопления, а также особые лимфатические сосуды - синусы, располагающиеся под капсулой и по ходу трабекул (рис. 2-4).

Рис. 2-4. Лимфатический узел. К - капсула, Т - трабекула, ПЛС - приносящий лимфатический сосуд, НК - наружная кора, ГК - глубокая кора, МВ - мозговое вещество. ЛУ - лимфатический узелок: К - корона, ГЦ - герминативный центр, СЗ - светлая зона центра, ТЗ - темная зона центра, МТ - мозговые тяжи, СКС - субкапсуярный синус, ПС - промежуточный синус.
Лимфатический узелок (фолликул) представляет собой сферическое скопление лимфоидной ткани, наружную границу которого образует слой уплощенных ретикулярных клеток. Различают первичные и вторичные узелки.
Первичные узелки - компактные однородные скопления малых (В) лимфоцитов рециркулирующего пула, связанных с ретикулярными клетками и особым видом антиген-представляющих фолликулярно-дендритных клеток.
Имеется небольшое количество Т-клеток, макрофагов. Встречаются в лимфатических узлах лишь в отсутствие антигенных воздействии (во внутриутробном периоде). Под влиянием антигенов превращаются во вторичные.
Вторичные узелки состоят из короны и герминативного центра.
Корона - скопление малых лимфоцитов на периферии узелка, полулунной формы на субкапсулярном полюсе и истончающееся до нескольких клеток на мозговом. Содержит В-клетки рециркулирующего пула и В-клетки памяти, а также незрелые плазматические клетки, мигрирующие из герминативного центра.
Герминативный центр развивается только под влиянием антигенной стимуляции вследствие Т-зависимого процесса. В нем происходит пролиферация и дифференцировка В-клеток в незрелые плазматические и В- клетки памяти в результате их взаимодействия с антигеном, фолликулярнодендритными клетками, Т-лимфоцитами (хелперами и супрессорами). Часть клеток, оказавшаяся неспособной к этим взаимодействиям, подвергается апоптозу и захватывается макрофагами. Из центра через глубокую кору в мозговые тяжи мигрируют незрелые плазматические клетки.
В разгар реакции на антиген герминативный центр включает темную зону (смежную с глубокой корой), и светлую зону (между темной и короной), содержащие делящиеся и дифференцирующиеся клетки. В светлей зоне рыхло располагаются более зрелые клетки, мигрировавшие в нее из темной, где они плотно прилежат друг к другу.
Межузелковая зона содержит малые лимфоциты и макрофаги; при антигенной стимуляции она почти полностью исчезает, замещаясь узелками.
Глубокая кора (паракортикальная зона) - Т-зависимая зона лимфатического узла. В ней осуществляются дозревание Т-клеток, поступивших из тимуса, а также их антиген-зависимая пролиферация и дифференцировка с формированием различных субпопуляций. Образована диффузной лимфоидной тканью, представленной Т-клетками, лежащими в петлях ретикулярной ткани и взаимодействующими с особым видом антиген-представляющих клеток - интердигитирующими клетками. Последние обладают цитоплазматическими отростками, охватывающими лимфоциты и проникающими между отростками соседних клеток. Встречаются также плазматические клетки, мигрирующие из узелков в мозговое вещество. Имеются лимфатические синусы
(промежуточные) и посткапиллярные венулы с высоким эндотелием, который способен взаимодействовать с хоминг-рецепторами Т- и В-лимфоцитов, обусловливая их миграцию из сосудистого русла.
Мозговое вещество образовано ветвящимися в анастомозирующими тяжами лимфоидной тони (мозговыми тяжами), между которыми располагаются соединительнотканные трабекулы и мозговые лимфатические синусы.
Мозговые тяжи являются В-зависимой зоной и содержат многочисленные плазматические клетки и (в менышем числе) В-лимфоциты и макрофаги. Плазматические клетки могут длительно находиться в тяжах и активно секретировать антитела в лимфу или поступать в нее, а далее - в кровоток.
10.Строение лимфатического узла. Источники развития.
Строение. Лимфатические узлы располагаются по ходу кровеносных и лимфатических сосудов, имеют овальную или бобовидную форму. Снаружи лимфатические узлы покрыты соединительнотканной капсулой, от которой отходят трабекулы. Капсула и трабекулы включают коллагеновые и эласти-ческие волокна и гладкие миоциты. На вогнутой поверхности лимфатических узлов находятся ворота. В ворота входят ар¬терия и нервы, выходят вены и выносящий лимфатический сосуд. Приносящие лимфатические сосуды входят с противо¬положной выпуклой стороны. Стромой лимфатических узлов является ретикулярная ткань, состоящая из ретикулярных клеток и ретикулярных волокон. Диаметр лимфатических уз¬лов от 0,5 до 1 см.
По периферии лимфатических узлов расположено более темное корковое вещество, представленное лимфатическими узелками, а в центре — более светлое мозговое вещество, представленное мозговыми тяжами. В лимфатических узлах имеются кортикальная зона (корковое вещество), мозговое вещество и паракортикальная зона, расположенная между кортикальной зоной (лимфатическими узелками) и тяжами мозгового вещества.
Источники развития:
1) мезенхима – с/тканная строма, кровеносные сосуды
2) СКК – лимфоидная паренхима
3) нейроэктодерма.
Функции:
1) лимфоцитопоэз,
2) участие в иммунной защите,
3) барьерная,
4) депонирование крови и лимфы,
5) очищение лимфы
11.Строение и функция лимфоидного узелка.
Лимфатический узелок (фолликул) представляет собой сферическое скопление лимфоидной ткани, наружную границу которого образует слой уплощенных ретикулярных клеток. Различают первичные и вторичные узелки.
Первичные узелки - компактные однородные скопления малых (В) лимфоцитов рециркулирующего пула, связанных с ретикулярными клетками и особым видом антиген-представляющих фолликулярно-дендритных клеток.
Имеется небольшое количество Т-клеток, макрофагов. Встречаются в лимфатических узлах лишь в отсутствие антигенных воздействии (во внутриутробном периоде). Под влиянием антигенов превращаются во вторичные.
Вторичные узелки состоят из короны и герминативного центра.
+
Корона - скопление малых лимфоцитов на периферии узелка, полулунной формы на субкапсулярном полюсе и истончающееся до нескольких клеток на мозговом. Содержит В-клетки рециркулирующего пула и В-клетки памяти, а также незрелые плазматические клетки, мигрирующие из герминативного центра.
Герминативный центр развивается только под влиянием антигенной стимуляции вследствие Т-зависимого процесса. В нем происходит пролиферация и дифференцировка В-клеток в незрелые плазматические и В- клетки памяти в результате их взаимодействия с антигеном, фолликулярнодендритными клетками, Т-лимфоцитами (хелперами и супрессорами). Часть клеток, оказавшаяся неспособной к этим взаимодействиям, подвергается апоптозу и захватывается макрофагами. Из центра через глубокую кору в мозговые тяжи мигрируют незрелые плазматические клетки.
В разгар реакции на антиген герминативный центр включает темную зону (смежную с глубокой корой), и светлую зону (между темной и короной), содержащие делящиеся и дифференцирующиеся клетки. В светлей зоне рыхло располагаются более зрелые клетки, мигрировавшие в нее из темной, где они плотно прилежат друг к другу.
Межузелковая зона содержит малые лимфоциты и макрофаги; при антигенной стимуляции она почти полностью исчезает, замещаясь узелками.
Глубокая кора (паракортикальная зона) - Т-зависимая зона лимфатического узла. В ней осуществляются дозревание Т-клеток, поступивших из тимуса, а также их антиген-зависимая пролиферация и дифференцировка с формированием различных субпопуляций. Образована диффузной лимфоидной тканью, представленной Т-клетками, лежащими в петлях ретикулярной ткани и взаимодействующими с особым видом антиген-представляющих клеток - интердигитирующими клетками. Последние обладают цитоплазматическими отростками, охватывающими лимфоциты и проникающими между отростками соседних клеток. Встречаются также плазматические клетки, мигрирующие из узелков в мозговое вещество. Имеются лимфатические синусы
(промежуточные) и посткапиллярные венулы с высоким эндотелием, который способен взаимодействовать с хоминг-рецепторами Т- и В-лимфоцитов, обусловливая их миграцию из сосудистого русла.
12.Глубокая кора. Клеточный состав. Функция.
Глубокая кора (паракортикальная зона) - Т-зависимая зона лимфатического узла. В ней осуществляются дозревание Т-клеток, поступивших из тимуса, а также их антиген-зависимая пролиферация и дифференцировка с формированием различных субпопуляций. Образована диффузной лимфоидной тканью, представленной Т-клетками, лежащими в петлях ретикулярной ткани и взаимодействующими с особым видом антиген-представляющих клеток - интердигитирующими клетками. Последние обладают цитоплазматическими отростками, охватывающими лимфоциты и проникающими между отростками соседних клеток. Встречаются также плазматические клетки, мигрирующие из узелков в мозговое вещество. Имеются лимфатические синусы (промежуточные) и посткапиллярные венулы с высоким эндотелием, который способен взаимодействовать с хоминг-рецепторами Т- и В-лимфоцитов, обусловливая их миграцию из сосудистого русла.
13.Строение мозгового вещества лимфатического узла.
Мозговое вещество образовано ветвящимися анастомозирующими тяжами лимфоидной тони (мозговыми тяжами), между которыми располагаются соединительнотканные трабекулы и мозговые лимфатические синусы.
Мозговые тяжи являются В-зависимой зоной и содержат многочисленные плазматические клетки и (в менышем числе) В-лимфоциты и макрофаги. Плазматические клетки могут длительно находиться в тяжах и активно секретировать антитела в лимфу или поступать в нее, а далее - в кровоток.
14.Т- и В- зависимые зоны лимфатического узла.
То же что и в 9 вопросе
15.Синусы лимфатического узла.
Лимфатические синусы - система особых внутриорганных лимфатических сосудов в корковом и мозговом веществе, обеспечивающая медленный ток лимфы через узел, в процессе которого она очищается (примерно на 99%) от содержащиеся в ней частиц (с извлечением антигенного материала) и обогащается антителами, клетками лимфоидного ряда и макрофагами.
Направление лимфотока в лимфатическом узле: из приносящих сосудов лимфа последовательно попадает в субкапсулярный, промежуточный и мозговой синусы, имеющие сходное строение, откуда (в воротах узла) направляется в выносящие сосуды.
Субкапсулярный (краевой, маргинальный) синус служит первым фильтрационным барьером для лимфы, попадающей в лимфатический узел. Он занимает щелевидное пространство между капсулой узла и наружной корой.
Выстлан особыми плоскими береговыми клетками, которые одни исследователи считают эндотелиальными, в другие - уплощенными ретикулярными. Эта выстилка лишена базальной мембраны, непрерывна со стороны капсулы, и прерывиста со стороны узелков, с многочисленными межклеточными щелями и подлежащим слоем маргинальных макрофагов. В просвете синуса находится сеть отростчатых ретикулярных клеток и волокон (замедляющая ток лимфы) с фиксированными на них и блуждающими макрофагами; имеются также многочисленные лимфоциты, плазматические клетки.
Промежуточный синус служит продолжением субкапсулярного и располагается между трабекулами и лимфоидной тканью наружной и глубокой коры.
Мозговой синус продолжает промежуточный и лежит между трабекулами и мозговыми тяжами.
Клиническое значение лимфатических узлов обусловлено не только их непосредственной ролью в развитии иммунных реакций, но возможностью возникновения в них очагов инфекции (распространяющейся лимфогенно), а
также метастазов злокачественных опухолей. Множественное увеличение лимфатических узлов характерно для опухолей лимфоидной ткани.
16.Строение селезенки. Источники развития
Селезенка - периферический и самый крупный орган иммунной системы,
располагающийся по ходу кровеносных сосудов. К ее основный функциям относятся.
1)участие в формировании гуморального и клеточного иммунитета, задержка антигенов, циркулирующих в крови;
2)разрушение старых и поврежденных эритроцитов и тромбоцитов;
3)депонирование крови и накопление тромбоцитов (до 1/3 общего их числа в организме).
Покрыта брюшиной и капсулой из плотной соединительной ткани, содержащей гладкомышечные клетки. От капсулы вглубь органа отходят трабекулы, анастомозирующие друг с другом. Паренхима (пульпа) включает два отдела с разными функциями: белую и красную пульпу (рис. 2-5).

Рис. 2-5. Селезенка. М - мезотелий, К - капсула, Т – трабекула, ЛУ - лимфатический узелок, ПАЛВ - периартериальное лимфатическое влагалище, ВС - венозные синусы, СТ - селезеночные тяжи.
Белая пульпа (около 20% объема органа) представлена лимфоидной тканью, расположенной по ходу артерий, и включает (1) лимфатическим узелки, (2) периартериальные лимфатическим влагалища (ПАЛВ) и (3) маргинальную зону

(рис. 2-6). К ее функциям относят обеспечение: (а) улавливания из крови антигенов, (б) взаимодействия лимфоцитов с антигенами, антигенпредставляющими клетками и друг с другом, (в) начальных этапов антигензависимой пролиферации и дифференцировки.
У человека селезенка закладывается на 5-й неделе эмбрионального периода развития в толще мезенхимы дорсальной брыжейки. В начале развития селезенка представляет собой плотное скопление мезенхимных клеток, пронизанное первичными кровеносными сосудами. В дальнейшем часть клеток дифференцируется в ретикулярную ткань, которая заселяется стволовыми клетками.
Одновременно с развитием узелков происходит формирование красной пульпы, которая становится морфологически различимой на 6-м месяце внутриутробного развития.
(1) лимфатические узелки (фолликулы, мальпигиевы тельце) располагаются по периферии ПАЛВ и по своей структурной и функциональной организации сходны с аналогичными образованиями в лимфатических узлах. Являются В-зависимой зоной селезенки.
(2)периартериальные лимфатические влагалище (ПАЛВ) окружают центральные артерии, состоят из цилиндрических компактных скоплений лимфоидной ткани, содержащей лимфоциты, макрофаги, ретикулярные и антиген-представляющие интердигитирующие клетки. Являются Т-зависимой зоной селезенки.
(3)маргинальная зона располагается в виде тонкого слоя к периферии от ПАЛВ и узелков на границе белой и красной пульпы, рядом с маргинальным синусом и содержит лимфоциты (преимущественно В-клетки), ретикулярные клетки и макрофаги. В ее наружной части накапливаются незрелые плазматические клетки, мигрирующие в красную пульпу для дозревания. Служит местом начального поступления в белую пульпу селезенки Т- и В-клетокь(направляющихся в дальнейшем в соответствующие зоны) н антигенов, которые здесь захватываются макрофагами.
Красная пульпа (около 75% объема органа) включает (1) венозные синусы и
(2) селезеночные или пульпарные тяжи (Бильрота). К ее функциям относятся: (а) депонирование зрелых форменных элементов крови; (б) контроль состояния иразрушение старых и поврежденных эритроцитов и тромбоцитов; (в) фагоцитоз инородных частиц; (г) обеспечение дозревания лимфоидных клеток ипревращения моноцитов в макрофаги.
(1)венозные синусы - тонкостенные анастомозирующие сосуды диаметром 12-50 мкм неправильной формы, образующие основную часть красной пульпы.
Выстланы эндотелиальными клетками необычной веретеновиднойь(палочкообразной) формы с узкими (1-3 мкм) щелями между ними, через которые в просвет синусов из окружающих тяжей мигрируют форменные элементы. Снаружи эти клетки охвачены циркулярно идущими отростками ретикулярных клеток и ретикулярными волокнами; базальная мембрана имеется лишь в отдельных участках.
(2)селезеночные (пульпарные) тяжи (Бильрота) - скопления форменных элементов крови (эритроцитов, тромбоцитов, лейкоцитов), а также макрофагов и плазматических клеток, лежащие в петлях ретикулярной ткани между синусами, в просвет которых они постоянно мигрируют. Старые, патологически измененные или поврежденные форменные элементы, (в первую очередь, эритроциты) с измененными маркерами и неспособные к миграции в синус, целиком фагоцитируются и перевариваются макрофагами, которые в тяжах образуются из моноцитов. Усиленное разрушение эритроцитов в селезенке может приводить к развитию анемии.
17.Кровеносная система селезенки.
Кровообращение в селезенке обладает рядом особенностей, обеспечивающих выполнение ее функций. В ворота органа входит селезеночная артерия, ветви которой проникают в трабекулы (трабекулярные артерии) и далее - в пульпу (пульпарные артерии). В пульпе адвентиция такой артерии замешается оболочкой из лимфоидной ткани, и артерия получает название центральной. Центральная артерия - мелкая, мышечного типа, по мере прохождения в белой пульпе отдает коллатерали в виде капилляров, снабжающих лимфоидную ткань и заканчивающихся в маргинальной зоне. Дистально центральная артерия утрачивает лимфоидную оболочку и, проникая в красную пульпу, разветвляется на 2-6 кисточковых артериолы, переходящие в эллипсоидные (гильзовые) капилляры (окружены эллисоидом или гильзой из ретикулярной ткани, лимфоцитов и макрофагов). Они изливают кровь непосредственно в венозные синусы (закрытое кровообращение) или между ними - в тяжи красной пульпы (открытое кровообращение), откуда она попадает в венозные синусы и далее в пульпарные н трабекулярные вены, собирающиеся в селезеночную вену.
Соотношение объема крови, направляющейся в открытую и закрытую системы, зависит от видовой принадлежности и функционального состояния. Закрытое (быстрое) кровообращение обеспечивает транспорт крови и насыщение тканей кислородом, открытое (медленное) - депонирование форменных элементов крови, возможность их сортировки и отбора полноценных жизнеспособных форм, контакт макрофагов с форменными элементами и антигенами, внесосудистое дозревание лимфоидных клеток макрофагов.
18.Строение и функция белой пульпы селезенки.
Белая пульпа – совокупность лимфоидной ткани в адвентиции внутриорганных артерий селезенки. Включает лимфатические узелки и периартериальные влагалища.
Лимфатические узелки – сходны по строению с аналогичными образованиями в лимфатических узлах.
4 зоны:
1) периартериальная,
2) центр размножения (светлый центр),
3) мантийная зона,
4) маргинальная зона.
1. Периартериальная зона – вокруг центральной артерии (одна из ветвей селезеночной артерии, эксцентрично расположена); тимусзависимая.
1) АПК,
2) некомитированные Т-л,
3) Т-лимфобласты
4) Т-пролимфоциты
5) субклассы Т-л Отсюда комитированные Т-л мигрируют в краевую зону.
2. Центр размножения – В-зона
1) макрофаги - АПК
2) вторичные В-л/бласты,
3) плазмоциты,
4) В-л на промежуточных стадиях развития
5) В-л памяти
3. Мантийная зона – окружает центр размножения и периартериальную зону. Образована макрофагами, зрелыми (малыми) В-л, комитир. Т-л (киллеры ….) центр размножения и мантийная зона – тимус-независимая зона.
4. Маргинальная зона –
1) комитированные и некомитированные Т- и В-л,
2) макрофаги, В маргинальной зоне синусные капилляры – выход лимфоцитов в кровоток.
Периартериальные лимфатические влагалища – скопления В-л, Т-л, плазмоцитов по ходу пульпарной артерии.
ИЛИ, МОЖНО НАПИСАТЬ ПО-ДРУГОМУ
Белая пульпа — это совокупность лимфоидных фолликулов (узелков), лежащих вокруг центральных артерий. Белая пульпа составляет 1/5 часть селезенки. Лимфоидные узелки селезенки отличаются по строению от фолликулов лимфоузла, так как содержат и Т-зоны и В-зоны. Каждый фолликул имеет 4 зоны:
• реактивный центр (центр размножения);
• мантийная зона — корона из малых В-лимфоцитов памяти;
• маргинальная зона;
• периартериальная зона или периартериальная лимфоидная муфтазона вокруг центральных артерий.
1-я и 2-я зоны соответствуют лимфоидным узелкам лимфоузла и являются В-зоной селезенки. В центре размножения фолликулов располагаются фолликулярные дендритные клетки, В-лимфоциты на разных стадиях развития и делящиеся В-лимфоциты, претерпевшие бласттрансформацию. Здесь происходит бласттрансформация и размножение В-лимфоцитов. В мантийной зоне происходит кооперация Т- и В-лимфоцитов и накопление В-лимфоцитов памяти.
Т-лимфоциты, составляющие 60 % всех лимфоцитов белой пульпы, лежат вокруг центральной артерии в 4-й зоне, поэтому эта зона является Т-зоной селезенки. Снаружи от периартериальной и мантийной зон узелков находится маргинальная зона. Ее окружает маргинальный синус. В этой зоне происходят кооперативные взаимодействия Т- и В-лимфоцитов, через нее в белую пульпу поступают Т- и В-лимфоциты, а также антигены, которые здесь захватываются макрофагами. Через эту зону в красную пульпу мигрируют созревшие плазмоциты. Клеточный состав маргинальной зоны представлен лимфоцитами, макрофагами, ретикулярными клетками.
Т- и В-зависимые зоны в белой пульпе селезенки.
(2)периартериальные лимфатические влагалище (ПАЛВ) окружают центральные артерии, состоят из цилиндрических компактных скоплений лимфоидной ткани, содержащей лимфоциты, макрофаги, ретикулярные и антиген-представляющие интердигитирующие клетки. Являются Т-зависимой зоной селезенки.
(1) лимфатические узелки (фолликулы, мальпигиевы тельце) располагаются по периферии ПАЛВ и по своей структурной и функциональной организации сходны с аналогичными образованиями в лимфатических узлах. Являются В-зависимой зоной селезенки.
Строение и функции красной пульпы селезенки.
Красная пульпа (около 75% объема органа) включает (1) венозные синусы и (2) селезеночные или пульпарные тяжи (Бильрота). К ее функциям относятся: (а) депонирование зрелых форменных элементов крови; (б) контроль состояния и разрушение старых и поврежденных эритроцитов и тромбоцитов; (в) фагоцитоз инородных частиц; (г) обеспечение дозревания лимфоидных клеток и превращения моноцитов в макрофаги.
(1)венозные синусы - тонкостенные анастомозирующие сосуды диаметром 12-50 мкм неправильной формы, образующие основную часть красной пульпы. Выстланы эндотелиальными клетками необычной веретеновидной (палочкообразной) формы с узкими (1-3 мкм) щелями между ними, через которые в просвет синусов из окружающих тяжей мигрируют форменные элементы. Снаружи эти клетки охвачены циркулярно идущими отростками ретикулярных клеток и ретикулярными волокнами; базальная мембрана имеется лишь в отдельных участках.
(2)селезеночные (пульпарные) тяжи (Бильрота) - скопления форменных элементов крови (эритроцитов, тромбоцитов, лейкоцитов), а также макрофагов и плазматических клеток, лежащие в петлях ретикулярной ткани между синусами, в просвет которых они постоянно мигрируют. Старые, патологически измененные или поврежденные форменные элементы, (в первую очередь, эритроциты) с измененными маркерами и неспособные к миграции в синус, целиком фагоцитируются и перевариваются макрофагами, которые в тяжах образуются из моноцитов. Усиленное разрушение эритроцитов в селезенке может приводить к развитию анемии.
Особенности микроциркуляторного русла селезенки.
Селезенка имеет очень хорошее кровоснабжение и выполняет роль депо крови наряду с другими органами (печень, мышцы), поэтому она способна значительно изменяться в объеме. Функции селезенки заключаются в кровотворении, регуляции и обеспечении иммунной защиты от проникающих инородных тел белковой природы и других посторонних для организма веществ, частиц и микроорганизмов. В селезенке разрушаются старые (недееспособные) эритроциты. Сравнительно с величиной органа селезеночная артерия отличается крупным диаметром. Близ ворот она распадается на 3-4 ветви, входящих каждая отдельно в толщу органа. Артерия делится на трабекулярные артерии, переходящие в пульпарные сосуды. В пульпе адвентиция части артериол окружена лимфоидными фолликулами и влагалищами. Такая артериола называется центральной. Далее по ходу центральная артериола проникает в красную пульпу и ветвится на несколько кисточек, penicilli. Кисточковые артериолы имеют утолщенную стенку из-за хорошего развития гладких мышечных клеток и ретикулярной ткани. В дистальном конце артериола содержит утолщение – гильзу. В гильзе имеются сократительные элементы. Она хорошо развита у свиней и слабо – у жвачных животных. Артериолы открываются в колбочковидные выпячивания или эллипсоидные капилляры.
Артериальные капилляры переходят в венозные синусы, стенки которых образованы эндотелиальным синцитием с многочисленными щелями, через которые кровяные элементы и попадают в венозные синусы. Начинающиеся отсюда венозные стволики в отличие от артериальных образуют между собой многочисленные анастомозы. Корни селезеночной вены (вены 1-го порядка) выносят кровь из относительно изолированных участков паренхимы органа, называемых зонами селезенки.
Кровь от нее оттекает в воротную вену, разветвляющуюся в печени до микрососудов. Здесь происходит ее обработка клетками печени, после чего кровь выводится в нижнюю полую вену.
Открытый ток: артериальный ворот селезёнки > трабекулярные артерии > прекапилляры > кисточковые артерии (соединительнотканные гильзы при сокращении препятствуют току крови обратно из пульпы в сосуды артериальной системы) > колбочковидные расширения с отверстиями, через которые в пульпу поступает кровь. Венозная система начинается с синусов, снабжённых отверстиями. Селезёнка депонирует кровь, концентрирует её (60 % форменных элементов). Жидкость всасывается в лимфоидную ткань селезёнки. Опорожнение селезёнки происходит при сокращении капсулы.
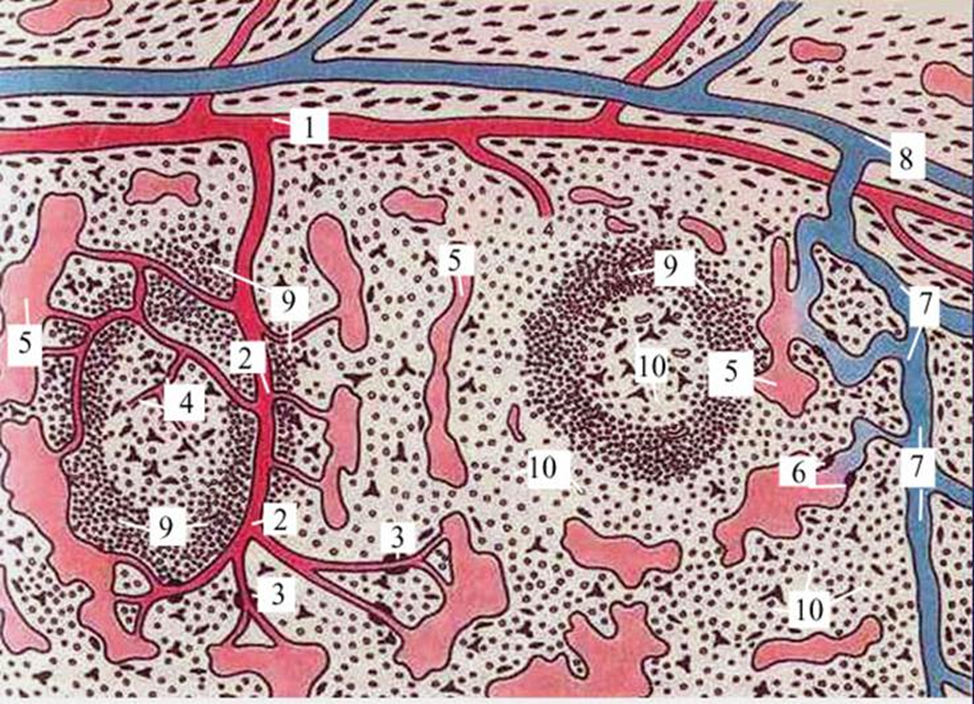
Схема: микроциркуляторное русло селезенки:
1 – трабекулярные артерии; 2 – прекапилляры; 3 – кисточковые артерии; 4 - 5 – колбочковидные расширения; 6 – посткапилляры;