

307_p344_D1_2301
.pdfА.Я. НИКИТИН, А.М. АНТОНОВА
УЧЕТЫ, ПРОГНОЗИРОВАНИЕ И РЕГУЛЯЦИЯ ЧИСЛЕННОСТИ ТАЕЖНОГО КЛЕЩА
В РЕКРЕАЦИОННОЙ ЗОНЕ ГОРОДА ИРКУТСКА
ИРКУТСК 2005
А.Я. Никитин, А.М. Антонова
Учеты, прогнозирование и регуляция численности таежного клеща в рекреационной зоне города Иркутска
Иркутск 2005
Рецензенты: доктор медицинских наук А.Д. Ботвинкин кандидат биологических наук О.В. Мельникова
Печатается по рекомендации ученого Совета НИИ биологии при Иркутском государственном университете
УДК 595.41.421:576.89 (571.53) ББК 28.691.8 (2 Рос – 4 Ирк)
Никитин А.Я., Антонова А.М. Учеты, прогнозирование и регуляция численности таежного клеща в рекреационной зоне города Иркутска. – Иркутск: Иркут. ун-т, 2005. – с.
В монографии рассматриваются способы увеличения информативности методов учетов численности имаго таежного клеща и эпидемиологического прогнозирования ситуации по «клещевым» инфекциям в рекреационной зоне города Иркутска. В многолетних временных рядах изменения обилия имаго, обращаемости населения в медицинские учреждения по поводу присасывания клещей и заболеваемости клещевым энцефалитом выявлены низко- и высокочастотные циклы, которые в последние двадцать лет накладываются на линейный тренд роста всех перечисленных показателей. Проанализирована связь динамики этих процессов с изменениями ряда биотических, абиотических и социальных факторов. Обсуждаются пути повышения эффективности тактических приемов полевой дезинсекции с учетом имеющегося ассортимента разрешенных к использованию низкоперсистентных химических пестицидов.
Монография предназначена для научных работников, аспирантов и студентов медицинских и биологических высших учебных заведений.
Ил. 10, Табл. 17, Библиогр. 155 назв.
ISBN
©Никитин А.Я., Антонова А.М., 2005
©Иркутский государственный университет, 2005
3
О Г Л А В Л Е Н И Е
ВВЕДЕНИЕ
ГЛАВА 1. ПРИРОДНЫЕ ОЧАГИ «КЛЕЩЕВЫХ» ИНФЕКЦИЙ В РЕКРЕАЦИОННОЙ ЗОНЕ ГОРОДА ИРКУТСКА
1.1.Природный очаг трансмиссивных заболеваний
1.2.Таежный клещ
1.3.«Клещевые» инфекции рекреационной зоны
ГЛАВА 2. УЧЕТЫ ЧИСЛЕННОСТИ ВЗРОСЛЫХ КЛЕЩЕЙ
2.1.Расчет доверительного интервала и стандартной ошибки при анализе обилия клещей
2.2.Заполнение пропусков во временных рядах
2.3.Смыкание рядов
2.4.Роль показателя «оценка активности нападения таежного клеща на население» при мониторинге численности клещей
ГЛАВА 3. ПРОГНОЗИЗМЕНЕНИЙЧИСЛЕННОСТИКЛЕЩЕЙ
3.1.Сезонная и многолетняя динамика численности клещей в рекреационной зоне г. Иркутска
3.2.Способы прогнозирования
Прогноз, основанный на связи обилия клещей с действием внешних факторов
Прогноз на основе ретроспективного анализа тенденций развития ряда
Факторы, осложняющие оценку ожидаемого обилия переносчика и уровня заболеваемости населения
ГЛАВА 4. РЕГУЛЯЦИЯ ЧИСЛЕННОСТИ КЛЕЩЕЙ НИЗКОПЕРСИСТЕНТНЫМИ ПЕСТИЦИДАМИ
ЗАКЛЮЧЕНИЕ
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
4
ВВЕДЕНИЕ
Рекреационная зона Иркутска находится в ландшафтно-эпидемиологическом районе с высокой степенью опасности заражения населения клещевым энцефалитом и иксодовыми клещевыми боррелиозами. Роль основного переносчика возбудителей этих заболеваний принадлежит таежному клещу (Ixodes persulcatus). Региональный мониторинг изменений численности клещей, их вирусофорности, заболеваемости населения инфекциями, передающимися при укусах имаго, проводится давно и обсуждению этих вопросов посвящено значительное число публикаций. Однако в силу того, что за последнее десятилетие эпидемиологическая обстановка по «клещевым» инфекциям не улучшилась, а напротив обострилась, считаем возможным провести собственный анализ некоторых методических аспектов процедур учета и прогнозирования обилия, регуляции численности клещей. Надеемся, что проделанная работа будет полезной для специалистов. Мы старались рассмотреть недостаточно обсуждаемые стороны общей для биологов, медиков и дезинфекторов проблемы – необходимости снижения региональной заболеваемости инфекциями, передающимися при укусах клещей. В то же время, по нашему замыслу книга должна быть доступна для студентов биологических и медицинских высших учебных заведений, от участия которых в решении рассматриваемых вопросов в конечном итоге зависит эпидемиологическое благополучие в будущем. Поэтому предлагаемое изложение не свободно от обсуждения общеизвестных фактов, классических формулировок и исторических ссылок.
В монографии рассмотрены методы учетов численности имаго таежного клеща. Предлагаются подходы к увеличению их информативности и оптимизации способов проведения эпизоотологического мониторинга (обследования) рекреационной зоны города в условиях ограниченного финансирования исследовательских и профилактических работ; приводятся способы прогноза временных рядов, наиболее адекватно отражающие изменения численности и активности нападения имаго клещей, динамику эпидемиологической ситуации в Иркутске по «клещевым» инфекциям; в динамике исследованных параметров выделены тренды, низко- и высокочастотные циклы, для которых обсуждается их связь с факторами различной природы; показано, что на пиках и спадах кривой динамики активности нападений имаго на население доминируют различные генерации клещей. Это явление может влиять на характер взаимоотношений переносчика с возбудителями трансмиссивных болезней, а значит быть причиной межгодовых различий в уровне заболеваемости населения «клещевыми» инфекциями; обсуждаются пути повышения эффективно-
5
сти тактических приемов полевой дезинсекции с учетом имеющегося ассортимента разрешенных к использованию низкоперсистентных химических пестицидов.
Воснову работы положен анализ материалов из отчетов паразитологического отделения ЦГСЭН города Иркутска об изменениях численности таежного клеща в рекреационной зоне, количестве «покусов» и заболеваемости населения «клещевыми» инфекциями, собранные почти за тридцать пять лет. Ввиду изучения количественных по природе признаков (численность, активность нападения, уровень заболеваемости и т.д.), статистика является главным аналитическим инструментом исследования. Использованный математический аппарат включает различные методы, в том числе анализа и прогноза временных рядов, реализованные с помощью прикладных компьютерных программ Excel и Statistica [Боровиков, Ивченко, 2000; Дуброва, 2003; Никитин, Сосунова, 2003].
Впервой главе приведены общие сведения, необходимые читателю, незнакомому с проблемой, для лучшего понимания обсуждаемых вопросов. В последующих главах излагаются собственно результаты анализа материалов, проведенного авторами. При этом цитируемые в монографии источники составляют лишь незначительную часть имеющихся по проблеме работ, и отобраны в значительной мере субъективно, что делает необходимым принести наши извинения неупомянутым авторам, так как роль многих из них в решении поставленных вопросов ничуть не меньше.
Ко времени подведения итогов по ситуации с «клещевыми» инфекциями в рекреационной зоне города Иркутска 2004 г. работа над рукописью книги была завершена. Поэтому эти данные включены лишь в часть приводимых таблиц и уравнений.
Авторы выражают свою признательность за практическую помощь в работе сотрудникам паразитологического отделения ЦГСЭН г. Иркутска, и, прежде всего энтомологу Г.А. Чистофоровой. Книга не была бы написана без инициативы профессора Иркутского государственного университета, д.б.н. Г.В. Гречаного. Ему, а также зав. кафедрой Иркутского медицинского университета, д.м.н. А.Д. Ботвинкину, научному сотруднику Иркутского противочумного института Сибири и Дальнего Востока, к.б.н. О.В. Мельниковой, доценту Иркутского государственного педагогического университета, к.б.н. И.А. Сосуновой, авторы выражают признательность за помощь в работе над рукописью.
6
ГЛАВА 1. ПРИРОДНЫЕ ОЧАГИ «КЛЕЩЕВЫХ» ИНФЕКЦИЙ В РЕКРЕАЦИОННОЙ ЗОНЕ ГОРОДА ИРКУТСКА
1.1. Природный очаг трансмиссивных заболеваний
Заболевания, распространение которых происходит путем переноса их возбудителей насекомыми и клещами, по предложению Е.Н. Павловского названы «трансмиссивными» болезнями. Существование трансмиссивных заболеваний поддерживается благодаря последовательному переходу возбудителя из организма хозяина-донора в организм переносчика и через его посредство в организм хозяина-реципиента. Всю сумму процессов, связанных с такими переходами, называют циркуляцией возбудителя, а территорию, на которой осуществляется процесс – природным очагом. В качестве патогенов, передающихся человеку при укусах членистоногих, выступают вирусы, риккетсии, бактерии и простейшие [Павловский, 1947; Беклемишев, 1970; Злобин, 1999]. К «классическим» трансмиссивным инфекциям относятся малярия, чума, туляремия, омская геморрагическая лихорадка и множество других заболеваний, в распространении которых участвуют членистоногие [Беклемишев, 1970]. Причем в зоне умеренного климата, расположенной между 40-600 северной широты и частично занимаемыми территориями России (Сибирь, Дальний Восток), Японии, Северной Кореи, Китая и Монголии наблюдается преобладание «клещевых» природно-очаговых трансмиссивных инфекций. В отличие от этого в субтропической и тропической Азии доминируют возбудители с «комариным» механизмом передачи [Злобин, 1999].
Различают облигатно-трансмиссивные заболевания, возбудители которых циркулируют исключительно благодаря переносчикам, и заболевания факультативнотрансмиссивные, для которых этот путь – лишь один из возможных. В обоих случаях природные очаги трансмиссивных инфекций характеризуются отсутствием их функциональной связи с человеком [Павловский, 1947; Беклемишев, 1970]. Заражение людей, оказавшихся на территории природных очагов трансмиссивных болезней в контакте с члени- стоногими-переносчиками патогенов, происходит случайно. Границы природного очага формируются путем распространения популяции возбудителя, которое зависит от пространственной структуры накладывающихся друг на друга ареалов популяций его носителей и переносчиков, а также действия лимитирующих абиотических факторов [Беклемишев, 1970; Литвин, Коренберг, 1999; Вержуцкий, 1999].
С течением времени в результате хозяйственной деятельности человека и антропогенного изменения окружающей среды, происходит либо «затухание» очагов в результате разрушения паразитарных систем, вплоть до их исчезновения, либо животные-носители и
7
членистоногие-переносчики инфекции приспосабливаются к существованию в измененных условиях [Окулова, 1986].
Весенне-летний клещевой энцефалит явился именно той инфекцией, на базе исследования которой создано учение Е.Н. Павловского о природной очаговости. Для функционирования природных очагов клещевого энцефалита необходимо наличие паразитарной триады: возбудителя, переносчика и животных-доноров, которые являются сочленами биоценоза определенных биотопов. Кровососущие клещи и позвоночные животные – их прокормители, обеспечивают хранение и постоянную циркуляцию вируса на территории природных очагов. В качестве основного переносчика вируса во всех известных очагах клещевого энцефалита выступают виды группы persulcatus [Таежный клещ..., 1985]. Два вида из этой группы имеют наиболее обширные ареалы и называются лесным (I. ricinus) и таежным (I. persulcatus) клещами. Из них в окрестностях города Иркутска встречается только таежный клещ, формирующий один из 60 известных для территории бывшего
СССР комплексов региональных популяций вида. Таежный клещ в силу своих этологических, эколого-географических и физиологических особенностей способен сохранять и поддерживать циркуляцию вируса клещевого энцефалита, а также осуществлять широкомасштабный контакт с человеком, тем самым, приобретая существенное эпидемиологическое значение.
1.2. Таежный клещ
Первое описание таежного клеща дано в 1930 г. [Таежный клещ..., 1985]. Основы периодизации его жизненного цикла наиболее полно изложены в работах Ю.С. Балашова [1967; Таежный клещ ..., 1985 и др.]. Иксодовые клещи – облигатные временные эктопаразиты позвоночных животных, которые характеризуются сложным циклом развития, включающим яйцо и три активные фазы: личинку, нимфу, имаго. Каждая активная фаза развития – паразитическая, один раз в жизни питающаяся кровью млекопитающих или птиц и, реже, – рептилий.
Паразитирует таежный клещ по треххозяинному типу, то есть каждая стадия развития питается на новом прокормителе. Трофические связи I. persulcatus с позвоночными животными очень широкие и разнохарактерные. Насчитывают до 300 видов животных, на которых питается таежный клещ. Чтобы избежать чрезмерного давления паразита на их хозяев, каждая стадия имеет собственный сезонный максимум активности питания, а также предпочитаемый круг прокормителей. Кроме того, как отмечено в работе Ю.С. Короткова и Г.С. Кисленко [2002], этому же служит растянутый во времени выход клещей после зимней диапаузы. Личинки и нимфы питаются, главным образом, на мелких мышевидных и насекомоядных в ию-
8
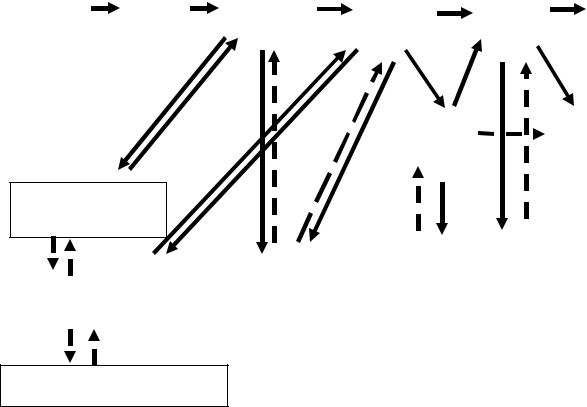
не-июле, а взрослые клещи – преимущественно на более крупных животных (зайцах, лисах, диких копытных) со второй декады мая по вторую декаду июня. Вблизи населенных пунктов половозрелые особи часто нападают на домашний скот и собак, что увеличивает вероятность контакта клещейсместным населением. Птицымогутбытьобъектомнападения всехтрехфаз развития членистоногих [Шихарбеев, 1965; Шихарбеев и др., 1970]. Следствием полигостальности клещей является высокая вероятность их распространения на новые территории, причем за счет птиц на более удаленные, чем обеспечивают млекопитающие [Львов, 1974; Schulze et al., 1986]. Однако без млекопитающих невозможно ни существование клещей, ни циркуляция вируса в очаге клещевого энцефалита [Павловский, 1947; Беклемишев, 1970; Наумов 1983; Гутова и др., 1985; Болотин, 2004]. Значительная роль в формировании природных очагов «клещевых» инфекций отводится механизмам трансфазовой и трансовариальной передачи вирусов и бактерий [Балашов, 1967; Расницин, 1976; Таежный клещ …, 1985; Балашов и др., 1998]. Принципиальная схема циркуляции вируса клещевого энцефалита, взятая нами с некоторыми изменениями и дополнениями из работ Р.Л. Наумова [1983; Таежный клещ …, 1985], приведенанарис. 1.1.
Особенностибиологии эктопаразита иегопрокормителей, характерихвзаимодействияс возбудителем клещевого энцефалита позволяют полагать, что на неэндемичных территориях, заселенных клещами, новые природные очаги вряд ли возникают вследствие заноса вируса носителями, особенно птицами. Это связано с редкостью межпопуляционных миграций у млекопитающих (исключая случаи их массовых переселений при вспышках численности), отсутствиему птицуровнявирусемиидостаточногодлязаражения новыхчленистоногих. Основной путь интродукции вируса и бактерий на новые (неэндемичные) территории – расселение (разнос) уже зараженных клещей, причем, в силу особенностей паразитирования, – преимущест-
веннонимфптицами[Кисленко, Коротков, 2002; Schulze et al., 1986; Smith et al., 1999].
Популяция таежного клеща представлена в течение теплого сезона года всеми фазами развития: имаго, нимфами, личинками, а также яйцами. Прямых методов определения абсолютного возраста отдельных особей (т.е. числа дней, прожитых ими от момента вылупления из яиц) в настоящее время нет. Большинство авторов считает, что основная часть популяции таежного клеща имеет трех- и четырехгодичный цикл развития с потенциально возможным диапазоном вариации от 2 до 6 лет, зависящим от условий обитания [Павловский, 1947; Балашов, 1967; Таежный клещ…, 1985; Валицкаяидр., 2001; Коротков, Кисленко, 2002].
9
Первое поколение |
Жизненный цикл таежного клеща |
Второе поколение |
||||||||
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
1 |
|
2 |
|
3 |
|
4 |
|
1 |
|
2 |
Имаго |
|
Яйцо |
|
Личинка |
|
Ним- |
|
Имаго |
|
Яйцо |
(самка) |
|
|
|
|
|
фа |
|
(самка) |
|
|
5 |
|
10 |
Имаго |
|
Человек |
(самец) |
|
|
6
Мелкие
млекопитающие
6 |
|
7 |
|
8 |
Мелкие |
|
Птицы, рептилии |
|
Крупные позвоночные |
млекопитающие |
|
|
|
|
9
Гамазовые клещи и блохи
Рис. 1. Схема паразитарной триады, обеспечивающей циркуляцию и сохранение вируса клещевого энцефалита в природном очаге. Сплошные лини – доказанные пути передачи вируса; штриховые – предполагаемые. (С изменениями по: [Наумов, 1983; Таежный клещ …, 1985]).
Пояснения. По трансфазовой (2–3–4–1 и 2–3–4–5) и трансовариальной (1–2) цепям вирус передается с большими потерями, которые увеличиваются при удлинении сроков развития переносчика, с повышением температуры и понижением влажности. Причем в процессе пассирования через организм клещей вирулентность вируса понижается [Чунихин и др., 1986].
Ведущая роль в прокормлении личинок и нимф, а также их заражении принадлежит мелким млекопитающим (3–6 и 4–6), особенно грызунам. Насекомоядные (бурозубки), часто являясь прокормителями клещей, не обеспечивают циркуляцию вируса, но могут быть фактором его сохранения в зимний период [Бахвалова и др., 2001]. Авторы расходятся во мнении при оценке роли прокормителей имаго в цепях 1–8 и 5–8 [Наумов и др., 1983].
Вопрос о значении птиц, рептилий в качестве доноров вируса (7–3 и 7–4) до конца не ясен. Их роль как прокормителей возрастает при избыточной численности переносчика в период популяционной депрессии мелких млекопитающих. Однако в лабораторных опытах у птиц не получены титры вирусемии, необходимые для заражения клещей. Участие рептилий и, прежде всего, ящериц в поддержании циркуляции вируса считалось вполне вероятным [Гутова и др., 1985], однако позже возможность присасывания отмечена только для I. ricinus I. pacificus [Григорьева, 2002].
Значение гамазовых клещей и блох в заражении мелких млекопитающих вирусом (6–9) до конца не ясно, но их участие в его циркуляции доказано [Земская, Пчелкина, 1974; Наумов, Гутова, 1984; Верета, Волков, 1986].
Известно, что самцы клещей способны заразить человека вирусом (5–10). Возможно, что укусы самцов увеличивают иммунную прослойку. Самки могут заражаться от самцов при копуляции (5-1), кроме того, особи одного и разных видов клещей передают вирус друг другу при совместном питании на одном прокормителе [Алексеев, 1990; Алексеев, Чунихин, 1991].
Для мелких млекопитающих показана возможность трансплацентарной (6–6) передачи возбудителя [Molnarova, Mayer, 1980], что несколько меняет наши представления о клещевом энцефалите, как облигатно трансмиссивной инфекции.