

1. Устройство и правила работы в бактериологической лаборатории.
Регламентация условий работы с возбудителями инфекционных заболеваний произведена в соответствии со степенью опасности микроорганизмов для человека. По этому признаку выделено четыре группы патогенных биологических агентов (ПБА):
Группа I: возбудители особо опасных инфекций (чума, натуральная оспа и др.)
Группа II: возбудители высококонтагиозных бактериальных, грибковых и вирусных инфекций (сибирская язва, холера, бешенство и др.)
Группа III: возбудители бактериальных, грибковых, вирусных и протозойных инфекций, выделенных в самостоятельные нозологические формы (коклюш, столбняк, туберкулез и др.)
Группа IV: возбудители бактериальных, грибковых, вирусных септицемий, менингитов, пневмоний, энтеритов, токсикоинфекций, острых отравлений (синегнойная инфекция и др.).
Большая часть микробиологических лабораторий работает с ПБА III и IV групп, а изучением возбудителей особо опасных инфекций (I и II группы) занимаются только специализированные лаборатории.
Базовые лаборатории, работающие с ПБА III и IV групп, должны соответствовать ряду требований (отдельное здание или отдельный вход, наличие систем водо-, электроснабжения, отопление, вентиляция и др.) и иметь необходимый набор помещений в соответствии с производственной мощностью и номенклатурой выполняемых исследований. Каждая лаборатория должна иметь «чистую» и «грязную» зоны.
«Грязная» зона включает помещения для приема и регистрации материала, боксы и комнаты для проведения микробиологических исследований, термостатная, автоклавная для обеззараживания материала. Окна и двери всех помещений должны герметично закрываться. Помещения для работы с живыми микроорганизмами должны быть оборудованы бактерицидными лампами, или иметь боксы биологической безопасности. Приточно-вытяжная вентиляция «грязной» зоны должна быть оборудована фильтрами тонкой очистки выбрасываемого воздуха. Обязательна маркировка столов, автоклавов, емкостей с дезинфицирующими растворами, стеллажей для чистого и инфицированного материала. Лаборатория должна быть оснащена специальной мебелью, иметь гладкие поверхности пола и стен, устойчивые к действию моющих и дезинфицирующих средств.
«Чистая» зона включает помещения для предварительных работ (моечная, препараторская, комната для приготовления и розлива питательных сред и др.), комнату для работы с документацией, помещения с холодильниками для хранения питательных сред и диагностических препаратов, гардероб для верхней одежды, комнату отдыха. В «чистой» зоне возможна работа с неживыми ПБА (серологические, молекулярно-генетические, биохимические исследования).
Обеспечение безопасности работы с патогенными микроорганизмами включает два основных фактора: технический и человеческий. Технический фактор – это наличие необходимых для работы помещений «чистой» и «грязной» зоны, оборудования, защитных систем и т.д. Человеческий фактор – это правильность действий человека по обеспечению безопасности, уровень владения профессиональной техникой, знание возможных источников и механизмов заражения, соответствующая подготовка и тренировка.
Работа в учебной бактериологической лаборатории также сопряжена с двумя опасными факторами – микроорганизмами, являющимися возбудителями инфекционных заболеваний и открытым огнем, что требует соблюдение мер противоэпидемической и противопожарной безопасности. Студенты обязаны ознакомиться с правилами техники безопасности и строго их соблюдать:
Необходимо:
работать в медицинских халатах с длинным рукавом, медицинских шапочках и бахилах;
личные вещи хранить в специально отведённом месте, верхнюю одежду оставлять в гардеробе;
содержать лаборатории в чистоте и порядке, свободными от материалов, не относящихся непосредственно к работе;
каждый вид деятельности осуществляется в определенной зоне: работа с микроорганизмами - на специально оборудованном лабораторном столе, заполнение протоколов – на рабочем столе;
при попадании инфицированного материала на стол, пол и другие предметы немедленно сообщить преподавателю и произвести дезинфекцию;
инфицированные материалы должны быть помещены в прочные непромокаемые контейнеры или контейнеры с дезинфицирующим раствором, которые закрывают перед удалением из лаборатории.
во время проведения работы двери лаборатории должны держаться закрытыми.
по окончании работы руки тщательно вымыть, а при необходимости обработать дезинфицирующим раствором.
Категорически запрещается:
принимать пищу в лаборатории
набирать жидкость в пипетку ртом
зажигать одну спиртовку от другой
переносить горящую спиртовку
оставлять спиртовку горящей после окончания использования по назначению
оставлять на рабочем месте нефиксированные препараты, чашки Петри с посевами и другую посуду с инфекционным материалом
касаться руками исследуемого материала в засеянных чашках Петри
2. Мир микробов. Особенности строения про- и эукариотической клетки.
Все живые существа, обитающие на Земле, можно условно разделить на представителей макро- и микромира. К макромиру относятся все живые существа, видимые невооруженным глазом, а к микромиру – представители живого мира, находящиеся за пределами разрешающей способности нашего глаза и обнаруживаемые с помощью различных оптических и электронных приборов. Размеры отдельных представителей колеблются от 0,01-0,4 мкм или 10-400 нм (вирусы) до 10 мкм и более (бактерии, грибы, простейшие).
Согласно современной классификации органического мира микроорганизмы могут быть представлены как клеточными (бактерии, грибы, простейшие), так и доклеточными формами (вирусы, вироиды, прионы).
Клеточные формы обладают характерными признаками живых существ: воспроизведение себе подобных, метаболизм (питание и дыхание), способность к активному движению, росту и размножению. Среди клеточных форм выделяют три домена (империи):
Archae – к этому домену относятся древнейшие обитатели нашей планеты (архебактерии) – метанобразующие бактерии, экстремально галофильные бактерии (растут в присутствии 12-32% NaCl) и термоацидофильные бактерии (растут при 75-90 °С и низком рН). Их клетка имеет примитивное прокариотическое строение и клеточную стенку, лишенную пептидогликанового слоя (Mendosicutes). Среди архебактерий патогенных для человека видов не обнаружено.
Bacteria – к этому домену относятся три типа бактерий: эубактерии с тонкой клеточной стенкой (Gracilicutes) - грамотрицательные, эубактерии с толстой клеточной стенкой (Firmicutes) – грамположительные, эубактерии без клеточной стенки (Tenericutes).
Eukaria – к этому домену относятся микроскопические простейшие, грибы и сине-зеленые водоросли.
К доклеточным формам существования относят:
Вирусы – наименьшие по размерам агенты, имеющие геном, окруженный белковой оболочкой и суперкапсидом (оболочечные вирусы). Вирусы являются облигатными внутриклеточными паразитами, репродуцирующимися только в живых клетках. Внеклеточная форма вируса - вирион, а внутриклеточная – вирус.
Прионы (от англ. proteinaceous infectious particle, белковоподобная инфекционная частица) – белковые инфекционные агенты, приводящие к развитию летальных неврологических заболеваний (губчатых энцефалопатий: куру, болезнь Кройтцфельдта-Якоба и др.)
Вироиды – небольшие молекулы кольцевой суперспирализованной РНК, не содержащие белковой оболочки и вызывающие заболевания растений (таблица 1)
Таблица 1. Мир микробов
Доклеточные формы |
Клеточные формы |
||||
Вирусы
Прионы
Вироиды |
Домен "Bacteria" |
Домен "Archae" |
Домен "Eukarуa" |
||
Прокариоты |
Эукариоты |
||||
Бактерии с тонкой клеточной стенкой, грамотрицательные (Gracilicutes) |
Архебактерии (Mendosicutes) |
Простейшие (царство Animalia, подцарство Protozoa) Типы: Sarcomastigofora Apicomplexa Ciliophora Microspora |
Грибы (царство Fungi) Типы: Zygomycota Ascomycota Basidiomycota Deuteromycota или митоспоровые грибы |
||
Бактерии с толстой клеточной стенкой, грамположительные (Firmicutes) |
|||||
Бактерии без клеточной стенки (Tenericutes) |
|||||
Анализ различий строения про- и эукариотической клетки обычно проводят по трем основным параметрам:
1) ядерно-генетическая организация клетки
2) организация мембранных структур клетки
3) функциональная организация клетки (таблица 2).
Таблица 2. Сравнительная характеристика прокариотов (эубактерий) и эукариотов
Признак |
Прокариотическая клетка |
Эукариотическая клетка |
Размер Анаэробное дыхание Фиксация азота Мембранные структуры |
1-10 мкм Возможно Возможна Отсутствуют |
10-100 мкм Обычно отсутствует Невозможна Имеются |
Генетический материал |
||
Расположение Форма Внехромосомная ДНК Гистоны Тип деления |
Нет ядерной мембраны Кольцевая молекула ДНК Располагается в плазмидах Отсутствуют Бинарный |
Есть ядерная мембрана Хромосома Располагается в митохондриях Имеются Митотический |
Синтез белка |
||
Рибосомы Место синтеза |
70S (50S и 30S субъединицы) Рибосомы, свободно расположенные в цитоплазме |
80S (60S и 40S субъединицы) Рибосомы в составе эндоплазматического ретикулума |
Клеточная стенка* |
||
Структурные элементы
Стеролы |
Образована пептидогликаном
Отсутствуют |
Содержит хитин и целлюлозу
Имеются |
* у эукариотических клеток нет клеточной стенки, а есть ЦПМ |
||
3. Систематика и номенклатура микроорганизмов.
Систематика (таксономия) бактерий является одним из наиболее важных и сложных разделов микробиологии. Задачами систематики являются классификация, номенклатура и идентификация организмов.
Классификация – распределение множества организмов по группам (таксонам). Любая классификация живых организмов призвана показать степень сходства и предполагаемую эволюционную взаимосвязь (при этом более высокие категории – емкие и широкие, а более низкие – конкретны и ограничены). Идентификация устанавливает принадлежность микроорганизмов к определенному таксону на основании наличия конкретных признаков.
Образование и применение научных названий микроорганизмов регламентируют «Международный кодекс номенклатуры бактерий», «Международный кодекс ботанической номенклатуры» (грибы), «Международный кодекс зоологической номенклатуры» (простейшие) и решения Международного комитета по таксономии вирусов. Все изменения научных названий микроорганизмов возможны лишь решениями соответствующих международных конгрессов и постоянных комитетов по номенклатуре.
Определение бактерий до вида важно не только с позиции чисто познавательной, общебиологической, но и связано с решением ряда прикладных и научных задач. Особенно это важно для медицинской, ветеринарной и промышленной микробиологии, где действующими объектами являются микроорганизмы, и мельчайшие неточности в определении вида могут привести к нежелательным последствиям. В настоящее время в микробиологии приняты два различных подхода к систематике, обусловливающих существование двух систем классификации: филогенетической (естественной) и фенотипической (искусственной). В основу филогенетической классификации положена идея создания системы прокариот, объективно отражающей родственные отношения между разными группами бактерий и историю их эволюционного развития. Фенотипическая классификация преследует, в первую очередь, практические цели, заключающиеся в том, чтобы быстрее установить принадлежность микроорганизма к определенному таксону.
В классификации бактерий используют набор фенотипических признаков: морфологических, культуральных, физиологических, биохимических и антигенных. Морфологическая идентификация (описание морфологических признаков) включает определение формы, размеров клеток, их взаимное расположение, количество и расположение жгутиков, наличие капсулы, способность образовывать споры, тинкториальные свойства (окраска по методу Грама, связанная со строением клеточной стенки и др. методы окраски). Однако, только морфологических признаков для идентификации бактерий недостаточно. При описании культуральных свойств, которые проявляются при культивировании бактерий в различных условиях, отмечают особенности роста бактерий на плотной питательной среде (размер, цвет, форма, характер колоний) и в жидких питательных средах (образование осадка, пленки, взвеси, хлопьев и т. д.). Однако, и этих признаков недостаточно, так как у культур различных видов они могут проявляться сходным образом. К числу физиологических признаков относятся возможность использовать те или иные источники углерода и азота, потребность в факторах роста, тип энергетических процессов (аэробное и анаэробное дыхание, брожение), отношение к температуре, влажности, кислотности среды и другим факторам внешней среды. В основу биохимической идентификации положены разнообразные биохимические признаки бактерий, обусловленные наличием ферментов, образованием определенных продуктов метаболизма (кислоты, спирты, газы и др.), типом запасных веществ, химическим составом клеток и т. д. Серологические критерии систематики основаны на специфических реакциях взаимодействия антигенов (компоненты клеточных стенок, жгутиков, капсул, ДНК и токсинов) идентифицируемых микроорганизмов с антителами, содержащимися в диагностических сыворотках. Такие серологические реакции, как агглютинация, преципитация, связывание комплемента, иммунофлуоресценция, иммуноферментный и радиоиммунный анализ, позволяют легко и быстро проводить предварительную идентификацию микроорганизмов.
В современной систематике бактерий достигнуты успехи в создании филогенетической системы классификации, отражающей основные направления эволюционного развития, но сохраняют свое значение и искусственные фенотипические классификации, более удобные для идентификации микроорганизмов.
Наиболее приемлемой филогенетической системой классификации прокариот является система, основанная на сопоставлении последовательности нуклеотидов в 16S-рРНК. Эта система положена в основу 2-го издания многотомной энциклопедии прокариот Bergey’s Manual of Systematic Bacteriology (Руководство по систематике бактерий Берджи), первый том которой вышел в свет в 2001 г. В этом труде все прокариоты разделены на 26 филогенетических «ветвей» (групп) на основании строения их 16S-рРНК; 23 «ветви» представлены эубактериями, а три – архебактериями. Следует подчеркнуть, что большое количество этих филогенетических групп содержат виды прокариот, которые не выделены в виде чистых культур и поэтому еще детально не изучены. Для представителей данных видов известны в настоящее время только последовательности нуклеотидов в 16S-рРНК. Из 23 групп эубактерий две филогенетические группы представлены грамположительными бактериями, остальные группы – грамотрицательными.
Основные таксономические категории
Номенклатура – присвоение названий отдельным группам и видам микроорганизмов.
Для микроорганизмов приняты следующие таксономические категории (по нисходящей): Домен – Царство – Тип (Отдел) – Класс – Порядок – Семейство – Род – Вид.
Названия видов биномиальны (бинарны), то есть обозначаются двумя словами – названия рода и вида на латинском языке: первое определяет их принадлежность к конкретному роду, второе – к виду. Например, Escherichia coli (кишечная палочка), Staphylococcus aureus (золотистый стафилококк), Streptococcus pyogenes (гноеродный стрептококк).
В микробиологии существуют также более мелкие таксономические единицы: подвид (subspecies), разновидность. Подвиды могут различаться по физиологическим (биовар), морфологическим (морфовар), по антигенным (серовар) свойствам, а также среде обитания (эковар).
В таблице 3 представлены основные таксономические категории для C.tetani и T.pallidum.
Таблица 3. Основные таксономические категории
Таксономические категории |
||
Домен |
Bacteria |
Bacteria |
Царство |
Prokaryotae |
Prokaryotae |
Тип* |
Firmicutes |
Gracilicutes |
Класс |
Clostridia |
Spirohaetes |
Порядок |
Clostridiales |
Spirochetales |
Семейство |
Clostridiaceae |
Spirochetaceae |
Род |
Clostidium |
Treponema |
Вид |
Tetani |
pallidum |
* Для таксонов высшего ранга предпочтительно название "Тип" (Phylum), а не "Отдел" (Division) |
||
4. Морфология и ультраструктура бактериальной клетки.
Единственная мембранная система прокариот – ЦПМ, входящая в состав клеточной стенки.
Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с рибосомами и неоформленного ядра, называемого нуклеоидом. Могут быть дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили, включения. Некоторые бактерии способны образовывать споры для переживания неблагоприятных условий.
Рибосомы
Нуклеоид
Цитоплазматичекие
включения

Жгутик
Клеточная
стенка
Цитоплазматическая
мембрана
Цитоплазма
Капсула
Ворсинки
(пили) Мезосома


Рис. 1. Строение прокариотической клетки
Капсула. Клеточную оболочку многих бактерий окружает слой аморфного, гидрофильного вещества. Капсула состоит из полисахаридов, иногда из полипептидов; Основную роль в организации капсул играет ЦПМ. Выделяют микрокапсулы (выявляемые только при электронной микроскопии) и макрокапсулы (обнаруживают при световой микроскопии). У некоторых бактерий полимеры клеточной оболочки, выделяемые наружу, свободно располагаются вокруг нее, образуя слизистый слой. Слизь растворима в воде. Капсула и слизь предохраняют бактерии от повреждений, высыхания, фагоцитов, воздействия бактерицидных факторов, обеспечивают адгезию на различных субстратах, могут содержать запасы питательных веществ.
Клеточная стенка. Она выполняет ряд важных функций: защищает бактерии от внешних воздействий, придает им характерную форму, поддерживает постоянство внутренней среды, участвует в делении, на поверхности клеточной стенки находятся рецепторы для бактериофагов, через клеточную стенку осуществляется транспорт питательных веществ и выделение метаболитов. Это прочная, упругая структура. Опорным каркасом клеточной стенки бактерий является пептидогликан (муреин, мукопептид).
О-специфическая
цепь
Ядро ЛПС
Липид А
ЛПС
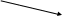

Периплазматическое пространство
Пептидогликан
Наружная
мембрана
Цитоплазмамическая мембрана







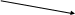








Грамположительные
бактерии
Грамотрицательные бактерии
Рис. 2. Строение клеточной стенки бактерий
Основным компонентом толстой клеточной стенки грамположительных бактерий является многослойный пептидогликан, составляющий 40-90 % массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты, молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. У большей части грамположительных бактерий также имеются периплазматические тейхоевые кислоты, располагающиеся между клеточной стенкой и ЦПМ, и являющиеся группоспецифическими антигенами. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков.
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами.
Цитоплазматическая мембрана. ЦПМ - это обязательная клеточная структура, являющаяся физическим, осмотическим, метаболическим барьером между внутренним содержимым бактериальной клетки и внешней средой. ЦПМ состоит из двух слоев фосфолипидов и встроенных в липидную мембрану белковых. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания, называемые мезосомами. Мезосомы образуют поперечные перегородки между делящимися клетками и являются местом прикрепления бактериальной хромосомы.
Цитоплазма. Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Ее основные составляющие – растворимые ферменты и РНК.
Бактериальный геном. Эквивалентом ядра у бактерий является нуклеоид (генофор). Он расположен в центральной зоне бактерий в виде двойной суперспирализованной кольцевой молекулы ДНК. Генофор не содержит гистонов. Кроме нуклеоида в бактериальной клетке имеются внехромосомные носители генетической информации – плазмиды, являющиеся ковалентно замкнутыми кольцами ДНК. Плазмиды несут ряд различных генов, кодирующих дополнительные (необязательные) признаки бактерий.
Рибосомы. Бактериальные рибосомы – сложные глобулярные образования, состоящие из молекул РНК и связанных с ними белками. Рибосомы необходимы для синтеза полипептидов. Диаметр рибосом составляет около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70S (единиц Сведберга). Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50S и 30S. Объединение субъединиц происходит перед началом трансляции.
Запасные гранулы. В цитоплазме бактерий содержатся различные включения, которые содержат временный избыток метаболитов. В виде гранул могут запасаться полисахариды, жиры, полимеры, полифосфаты (волютин), сера, кристаллизованные белки.
Жгутики. Жгутики бактерий являются органами движения бактерий. Расположение жгутиков – характерный признак, имеющий таксономическое значение. Жгутик – спирально изогнутая полая нить, образованная субъединицами белка флагеллина. Толщина жгутиков 12-20 нм, длина 3-15 мкм (больше длины клетки). Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду.
Микроворсинки. Поверхность многих бактерий покрыта цитоплазматическими выростами – микроворсинками, встречающимися у подвижных и неподвижных видов. Пили (фимбрии, ворсинки) — нитевидные образования, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина.
Споры. Споры образуются при неблагоприятных условиях существования бактерий. Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Способностью к спорообразованию обладают только бактерии родов Bacillus и Clostridium и многие сапрофиты. Высокая устойчивость спор связана с низким содержанием свободной воды, высокой концентрацией кальция, вялостью метаболических процессов, наличием дипиколиновой кислоты и белка, богатого цистеином, а также наличие нескольких оболочек, которые являются дополнительной защитой от неблагоприятных внешних воздействий. При попадании спор в благоприятные условия они прорастают.
5. Основные формы бактерий.
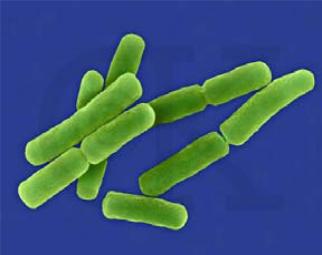
Отдельным видам бактерий с достаточным постоянством присущи определенная форма, размер и расположение. Длина бактериальных клеток варьирует от 0,1-0,2 мкм (виды Mycoplasma) до 10-15 мкм (виды Clostridium), толщина – от 0,1 до 2,5 мкм. Средние размеры бактерий 2-3х0,3-0,8 мкм. Выделяют три основные формы бактерий – шаровидные (кокки), палочковидные (цилиндрические) и извитые (спиральные).
 а)
а)
 б)
б)
 в)
в)
Рис. 3. Основные формы бактерий (микрофотографии)
а) шаровидные (кокки); б) палочковидные; в) извитые
Большинство кокков имеют шаровидную или овальную форму, клетки некоторых видов могут быть ланцетовидными (пневмококки) или бобовидными (нейссерии). По характеру расположения клеток в мазках выделяют диплококки (располагаются парами), стрептококки (располагаются цепочками), стафилококки (располагаются в виде гроздьев винограда), сарцины (располагаются пакетами по 8,16, 32 и более клеток) и др.
монобактерии диплобактерия стрептобактерия
Палочковидные
бактерии различаются по форме концов
клетки и взаимному расположению. Палочки
могут быть правильной (кишечная палочка),
неправильной (коринебактерии) формы,
ветвящиеся (актиномицеты). Палочковидные
бактерии могут располагаться в мазках
одиночно и беспорядочно (монобактерии),
попарно (диплобактерии) или цепочкой
(стрептобактерии, стрептобациллы)
(рис.4).

Диплококки
Стрептококки
Тетракокки
Стафилококки
Сарцины


Рис. 4. Основные формы бактерий
Извитые формы – представлены изогнутыми палочками. В зависимости от формы и количества завитков различают три типа клеток: вибрионы (от греч. vibrio - извиваюсь, изгибаюсь) имеют один завиток, не превышающий четверти оборота спирали (изогнутые клетки наподобие запятой); спириллы (от греч. speira – спираль) имеют 3–5 крупных завитков и спирохеты – большое количество мелких завитков (рис.5).
Вибрионы спириллы спирохеты
Рис. 5. Виды извитых клеток
6. Микроскопический метод диагностики инфекционных заболеваний.
Микроскопический метод диагностики инфекционных заболеваний включает приготовление мазка, его окраску с использованием простых и сложных методов, проведение иммерсионной микроскопии для изучения морфологических (формы и расположения) и тинкториальных свойств (отношение к красителю) с целью идентификации до рода (Staphylococcus, Streptococcus) или семейства (Enterobactericeae).
Первым этапом микроскопического метода является приготовление мазка (рис. 6). Мазок выполняют на чистом, обезжиренном предметном стекле, по поверхности которого капля воды должна растекаться равномерно. Для очистки стекла необходимо натереть его кусочком мыла и удалить мыло с помощью марлевой салфетки или протереть спиртом.
Для приготовления мазка на обратной стороне предметного стекла маркируют зону и наносят небольшую каплю воды или изотонического раствора хлорида натрия, в которой распределяют исследуемый материал. Если мазок готовят из жидкого исследуемого материала или микроорганизмов, культивированных в жидкой питательной среде, берут каплю жидкости петлей и равномерно распределяют в обозначенной зоне.
Препарат высушивают при комнатной температуре на воздухе. Хорошо приготовленный тонкий мазок высыхает равномерно и быстро. Если же высушивание препарата замедлено, то препарат подогревают, держа стекло мазком вверх, в потоке теплого воздуха высоко над пламенем горелки. Это нужно производить крайне осторожно, не допуская перегревания мазка, так как при этом может произойти слишком быстрое и грубое свертывание белков в протоплазме микробов, что нарушит их структуру. Если препарат высушен не полностью, то при фиксации он окажется также испорченным.
Фиксация препарата проводится с целью: 1) убить микробов и сделать безопасным дальнейшее обращение с ними; 2) прикрепить (фиксировать) мазок к стеклу, чтобы он не смывался при дальнейших манипуляциях жидкими красителями; 3) сделать микробов более восприимчивыми к окраске, там как убитые микробные клетки окрашиваются лучше, чем живые.
Способы фиксации. Существует два основных способа фиксации: физическая (термическая) и химическая (использование фиксаторов). В практической бактериологии обычно применяют фиксацию в пламени горелки (фиксация жаром) – метод грубый, но сохраняющий морфологию и отношение к красителям у бактерий. Для этого предметное стекло в положении мазком вверх 3 раза (в течение 3-5 сек) проводят через наиболее горячую часть пламени горелки. В ряде случаев используют жидкие фиксирующие растворы (этиловый и метиловый спирт, ацетон, формалин и др.), которые предотвращают ферментативный аутолиз бактерий и стабилизируют макромолекулы путем их химического сшивания, оказывая более щадящее действие. Для проведения химической фиксации препараты погружают в раствор фиксирующей жидкости на 5-20 минут. Выбор способа фиксации зависит от окрашиваемого препарата (жидкие фиксаторы используются для мазков крови, гноя и др.) и метода окраски.
1. приготовление 2.
высушивание 3. фиксация
4. окраска 5. Иммерсионная
микроскопия
мазка
микроскопия

Рис.6 Схема проведения микроскопического метода
7. Простые и сложные методы окраски бактерий.
Простой метод окраски. Является одноэтапным и заключается в окраске микропрепарата одним красителем. Используют основные анилиновые красители, такие как, фуксин, генцианвиолет, метиленовый синий в виде водных растворов или пропитанных красителем фильтровальных бумажек, которые помещают на мазок и смачивают водой. Продолжительность окраски составляет 1-5 минут, после чего микропрепарат промывают водой, высушивают и микроскопируют. В препаратах, окрашенных простым методом, можно получить представление о форме, расположении и размерах микробных клеток.
Сложные методы окраски. Сложные (дифференцирующие) методы окраски (по Граму, Цилю-Нильсену и др.) включают последовательное использование нескольких красителей и дополнительных способов обработки препаратов. Эти методы позволяют дифференцировать бактерии в зависимости от строения их структур, чаще всего поверхностных. Например, метод окраски по Граму позволяет дифференцировать бактерии по строению их клеточной стенки: грамположительные (фиолетовые) бактерии с толстой клеточной стенкой и грамотрицательные (красные) бактерии с тонкой клеточной стенкой.
Существуют специальные сложные методы окраски, которые используют для выявления структурных компонентов бактериальной клетки: капсул, цитоплазматических включений, спор, жгутиков.
8. Механизмы окрасок по Граму и Цилю-Нильсену.
Метод Грама. Относится к сложным методам окраски и позволяет дифференцировать грамположительные и грамотрицательные бактерии, что является важным таксономическим признаком. Результат окраски определяется особенностями строения клеточной стенки бактерий. При окраске генцианвиолетом и последующем воздействии раствора Люголя образуется комплексное соединение генцианвиолета и йода с пептидогликаном, которое при обработке спиртом удерживается в клетках грамположительных бактерий, имеющий многослойный пептидогликан и вымывается из грамотрицательных бактерий, имеющих тонкий слой пептидогликана. При дополнительной окраске водным раствором фуксина грамотрицательные бактерии приобретают красный цвет.
Таблица 4. Метод Грама
Этапы окраски |
Цвет бактерий |
|
Все бактерии окрашиваются в фиолетовый цвет
|
|
Все бактерии остаются фиолетовыми
|
|
Грамположительные бактерии остаются фиолетовыми, грамотрицательные обесцвечиваются |
|
Грамположительные остаются фиолетовыми, грамотрицательные окрашиваются в красный цвет |
Метод Циля-Нильсена. Относится к сложным методам окраски и позволяет дифференцировать кислотоустойчивые и некислотоустойчивые бактерии.
Таблица 5. Метод Циля-Нильсена
Этапы окраски |
Цвет бактерий |
|
Все бактерии окрашиваются в красный цвет
|
|
Все бактерии остаются красными |
|
Кислотоустойчивые бактерии остаются красными, некислотоустойчивые обесцвечиваются |
|
Кислотоустойчивые остаются красными, некислотоустойчивые окрашиваются в синий цвет |
Кислотоустойчивые бактерии имеют особое строение клеточной стенки, содержащей большое количество сложных липидов (миколовая и фтиоловая кислоты, воск D, глико- и фосфолипиды), что делает эти бактерии кислото-, щелоче- и спиртоустойчивыми. Клеточная стенка этих бактерий плохо воспринимает анилиновые красители и обычные способы окраски. Высокая температура в процессе окраски методом Циля-Нильсена расплавляет липиды, фенол разрыхляет клеточную стенку и краситель проникает внутрь клетки. После остывания препарата липиды вновь затвердевают, прочно удерживая краситель, поэтому кислотоустойчивые бактерии не обесцвечиваются серной кислотой и остаются красными. Некислотоустойчивые бактерии обесцвечиваются и их докрашивают контрастным красителем – метиленовым синим. В основном, метод используется для окраски бактерий, относящихся к роду Mycobacterium (M.tuberculosis, M.bovis, M.leprae и др.)
9. Специальные методы окраски для выявления отдельных структур бактериальной клетки (Бурри-Гинса, Ожешко, Леффлера, Нейссера).
Метод Бурри-Гинса. Относится к сложным методам окраски и используется для выявления капсулы бактериальной клетки.
Этапы окраски:
На предметном стекле смешать каплю взвеси микробных клеток с каплей туши, разведенной 1:10 и при помощи стекла со шлифованным краем сделать мазок таким же образом, как мазок крови, высушить и осторожно фиксировать в пламени горелки или горящим на мазке спиртом.
Нанести на мазок водный раствор фуксина на 1-2 мин.
Промыть водой, высушить на воздухе.
Метод Бурри-Гинса относится к негативным методам окраски: на первом этапе тушь, обтекая капсульные бактерии, создает черный или коричневый фон (соответствующий цвету туши) на котором хорошо видны неокрашенные бактерии, которые на втором этапе окрашиваются в красный цвет, а капсула остается бесцветной. Таким образом, капсулы бактерий хорошо видны на темном фоне туши виде неокрашенных зон вокруг красных бактерий.
Метод Ожешко (Ауэски). Относится к сложным методам окраски и используется для выявления спор бактерий.
Этапы окраски:
На нефиксированный мазок нанести 0,5% раствор (HCl) и подогреть в течение 2-3 мин на пламени (для протравливания плотной оболочки споры) до полного испарения кислоты. При этом происходит фиксация мазка к стеклу.
Окрасить по методу Циля-Нильсена.
Споры бактерий будут окрашиваться в красный цвет (как кислотоустойчивые бактерии), а вегетативные формы в синий цвет (как некислотоустойчивые бактерии).
Метод Леффлера. Относится к простым методам окраски и используется для выявления зерен волютина (цитоплазматических включений полифосфатов) у возбудителя дифтерии (C.diphtheriae), у которого эти включения располагаются по полюсам клетки. Фиксированный мазок окрашивают уксусно-кислым метиленовым синим в течение 3-5 минут. Зерна волютина обладают свойством метахромазии и более интенсивно накапливают краситель – цитоплазма бактерий окрашена в голубой цвет, зерна волютина – в темно-синиий.
Метод Нейссера. Относится к сложным методам и используется наряду с методом Леффлера для выявления зерен волютина.
На фиксированный мазок нанести ацетат синьки Нейссера (уксуснокислый раствор метиленового синего) на 1-2 мин.
Добавить раствор Люголя на 30 секунд и промыть водой.
Нанести водный раствор везувина или хризоидина на 1 минуту.
Зерна волютина имеют в отличие от цитоплазмы щелочную реакцию и избирательно воспринимают ацетат синьки, окрашиваясь в темно-синий цвет. Цитоплазма клетки, обладающая кислой реакцией, воспринимает щелочной краситель везувин и окрашивается в желтый цвет (метахромазия).
10. Методы окраски отдельных групп про- и эукариот (Романовского-Гимзы, Здродовского). Изучение подвижности микроорганизмов.
Для окраски отдельных групп микроорганизмов используют специальные методы окраски: Романовского-Гимзы, Здродовского и др.
Метод Романовского-Гимзы. Относится к сложным, универсальным методам окраски. Его применяют для обнаружения нуклеоида бактерий, а также окраски спирохет, хламидий, риккетсий, простейших и мазков крови.
Этапы окраски:
На мазок нанести рабочий раствор красителя (2 капли красителя на 1 мл дистиллированной воды), состоящего из смеси основного метиленового синего, азура и кислого эозина на 10-20 минут.
Препарат промыть и высушить.
ДНК нуклеоида окрашивается метиленовым синим и азуром в сине-фиолетовый цвет, цитоплазма клетки - эозином в розовый цвет. Представители разных родов спирохет окрашиваются в разные цвета по методу Романовского-Гимзы: боррелии – в сине-фиолетовый, трепонемы – бледно-розовый (T.pallidum – бледная трепонема), лептоспиры – красно-розовый.
Метод Здродовского используется для окраски риккетсий, являющихся облигатными внутриклеточными паразитами и вызывающими различные риккетсиозы. Этапы окраски:
Окрасить мазок разведенным фуксином Циля (10-15 капель на 10 мл дистиллированной воды) в течение 5 минут, промыть водой
Обработать мазок 0,5% раствором лимонной кислоты или 0,01% раствором хлористоводородной кислоты, промыть водой.
Окрасить метиленовым синим в течение 1 минуты.
Промыть водой и высушить препарат.
Риккетсии, окрашенные по методу Здродовского, имеют красный цвет, цитоплазма клеток, в которых они паразитируют – голубая, ядра – синие.
Для изучения подвижности микроорганизмов используют нативные препараты, приготовленные по методу «висячей» и «раздавленной капли».
Метод «висячей капли». Препарат готовят на покровном стекле, в центр которого наносят одну каплю исследуемого материала. Затем специальное предметное стекло с лункой, края которой предварительно смазывают вазелином, прижимают к покровному стеклу так, чтобы капля находилась в центре лунки. Быстрым движением переворачивают препарат покровным стеклом вверх. В правильно приготовленном препарате капля должна свободно висеть над лункой, не касаясь ее дна или края. Для микроскопии вначале используют объектив малого увеличения (8х или 10х), находят край капли, а затем устанавливают объектив 40х и исследуют препарат.
Метод «раздавленной» капли. На поверхность обезжиренного предметного стекла наносят каплю исследуемого материала или суспензию бактерий и покрывают ее покровным стеклом. Капля должна быть небольшой, не выходящей за край покровного стекла.
11. Виды микроскопии (фазово-контрастная, люминесцентная, темнопольная, электронная).
Иммерсионная микроскопия. Применяется для увеличения разрешающей способности метода световой микроскопии. Разрешающая способность системы светооптической микроскопии определяется длиной волны видимого света и числовой апертурой системы. Погружение объектива в среду (минеральное масло), имеющую высокий коэффициент преломления, близкий к таковому у стекла, препятствует рассеиванию света от объекта. Таким образом, достигается увеличение числовой апертуры и соответственно разрешающей способности. Для иммерсионной микроскопии применяют специальные иммерсионные объективы, маркированные черной полосой и снабженные меткой (МИ – масляная иммерсия).
Фазово-контрастная микроскопия. Предназначена для изучения нативных (живых и неокрашенных) препаратов за счет повышения их контрастности. При прохождении света через окрашенные объекты происходит изменение амплитуды световой волны, а при прохождении через неокрашенные объекты – фазы световой волны, что используют для получения высококонтрастного изображения. Для повышения контрастности фазовые кольца покрывают металлом, поглощающим прямой свет, не влияя на сдвиг фазы. В оптической системе микроскопа применяют специальный конденсор с револьвером диафрагм и центрирующим устройством. Неокрашенные объекты выглядят темными на светлом поле (позитивный фазовый контраст) или светлыми на темном фоне (негативный фазовый контраст).
Темнопольная микроскопия. Применяется для прижизненного изучения микробов в нативных неокрашенных препаратах. Микроскопия в темном поле зрения основана на явлении дифракции света при боковом освещении частиц, взвешенных в жидкости. Для этого используют темнопольный конденсор, выделяющий контрастирующие структуры неокрашенного материала. При этом способе освещения лучи от осветителя падают на объект сбоку, а в линзы микроскопа поступают только рассеянные лучи. В результате на темном фоне (неосвещенном поле зрения) видны ярко светящиеся частицы (микроорганизмы). С помощью темнопольной микроскопии изучают препараты типа «раздавленная капля». Предметные стекла должны быть не толще 1,1-1,2 мм, покровные — 0,17 мм, без царапин и загрязнений. При приготовлении препарата следует избегать наличия пузырьков и крупных частиц, поскольку эти дефекты будут видны ярко святящимися и не позволят наблюдать препарат.
Люминесцентная (флюоресцентная) микроскопия. Метод основан на явлении фотолюминесценции. Люминесценция (флюоресценция) – это способность некоторых объектов или веществ светиться при воздействии ультрафиолетового или другого коротковолнового излучения. При этом испускаемые световые волны длиннее волны, вызывающей свечение. Иными словами, флюоресцирующие объекты поглощают свет одной длины волны и излучают в другой области спектра. Например, если индуцирующее излучение синее, то образующееся свечение может быть красным или желтым. Различают первичную и вторичную люминесценцию. Первичная люминесценция (биолюминесценция) наблюдается без предварительного окрашивания за счет собственных люминесцирующих веществ, вторичная – возникает после окрашивания флюорохромами.
Люминесцентная микроскопия по сравнению с обычными методами обладает рядом преимуществ: возможностью исследовать живые микробы и обнаруживать их в исследуемом материале в небольших концентрациях вследствие высокой степени контрастности. Люминесцентная микроскопия нашла широкое применение для визуализации результатов иммунохимических реакций, основанных на специфическом взаимодействии меченых флюоресцирующими красителями антител с антигенами изучаемого объекта.
Электронная микроскопия. Для построения изображения в электронной микроскопии используют поток электронов, что позволяет изучить объекты, размеры которых лежат за пределами разрешающей способности светового микроскопа. В качестве «линз», фокусирующих электроны, служит электромагнитное поле, создаваемое электромагнитными катушками. Изображение в электронном микроскопе наблюдают на флюоресцирующем экране и фотографируют. Объекты при электронной микроскопии находятся в глубоком вакууме, поэтому подвергаются фиксации и специальной обработке. Кроме того, они должны быть очень тонкими, так как поток электронов сильно поглощается объектом. В связи с этим в качестве объектов используют ультратонкие срезы толщиной 20-50 нм, помещенные на тончайшие пленки. Разрешающая способность электронных микроскопов значительно выше, чем световых, и достигает 15 нм, что позволяет получить полезное увеличение в миллионы раз.
Наиболее широко используются два способа электронной микроскопии: просвечивающая (трансмиссивная) и сканирующая. Просвечивающая электронная микроскопия применяется для изучения ультратонких срезов микробов, тканей, а также строения мелких объектов (вирусов, жгутиков и др.), контрастированных фосфорно-вольфрамовой кислотой, уранилацетатом, напылением металлов в вакууме и др. Сканирующая электронная микроскопия применяется для получения трехмерного изображения поверхности исследуемого объекта.
12. Устройство биологического микроскопа. Порядок проведения иммерсионной микроскопии.
Для световой микроскопии используют различные виды микроскопов, которые подразделяются на студенческие, рабочие, лабораторные, исследовательские, различающиеся по конструкции и комплектации оптикой. Отечественные микроскопы («Биолам», «Бимам», «Микмед», МБР-1, МБИ-2 и др.) имеют обозначения, указывающие, к какой группе они относятся (С — студенческие, Р — рабочие, Л — лабораторные, И — исследовательские); комплектация обозначается цифрой.
Увеличение изображения достигают системой линз конденсора, объектива и окуляра. Конденсор, расположенный между источником света и изучаемым объектом, собирает лучи света в поле микроскопа. Объектив создает изображение поля микроскопа внутри тубуса. Окуляр увеличивает это изображение и делает возможным его восприятие глазом.
В микроскопе различают две части: механическую и оптическую. К механической части относятся: штатив (состоящий из основания и тубусодержателя), тубус с револьвером для крепления и смены объективов, предметный столик, механизмы для грубого (макровинт) и тонкого (микровинт) регулирования изображения. Оптическая часть микроскопа представлена объективами, окулярами и осветительной системой, которая, в свою очередь, состоит из расположенных под предметным столиком конденсора Аббе, зеркала, имеющего плоскую и вогнутую сторону, а также автономного или встроенного осветителя.
Порядок проведения иммерсионной микроскопии
Поднять конденсор микроскопа выше предметного стекла и полностью открыть диафрагму;
Включить осветитель и установить свет с помощью зеркала;
На мазок нанести каплю иммерсионного масла, поместить препарат на предметный столик микроскопа напротив объектива;
Поставить в рабочее положение иммерсионный объектив, маркированный черной линией (черным ободком);
Под контролем зрения, глядя сбоку, с помощью макровинта осторожно опустить тубус микроскопа до погружения иммерсионного объектива в каплю масла;
Глядя в окуляр, очень медленно опускать тубус при помощи макровинта по направлению к микропрепарату, пока не появится изображение объекта;
Далее провести точную фокусировку с помощью микровинта;
В процессе микроскопии следует медленно передвигать препарат, установить наилучшее поле зрения и изучить морфологические (форму и расположение) и тинкториальные (цвет) свойства микроорганизмов;
По окончании микроскопии поднять тубус и после этого снять препарат с предметного столика микроскопа;
Протереть фронтальную линзу иммерсионного объектива чистой марлевой салфеткой, смоченной спиртом, и повернуть револьвер таким образом, чтобы ни один из объективов не был направлен в сторону конденсора.
13. Особенности строения, размножения грибов, виды мицелия, спор.
Грибы — это эукариоты, относящиеся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) микроорганизмы с клеточной стенкой. Они широко распространены в природе, особенно в почве.
Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннанов, глюканов, целлюлозы, хитина), а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы (в отличие от холестерина — главного стерола тканей млекопитающих). Грибы являются грамположительными микробами, вегетативные клетки — некислотоустойчивые. Тело гриба называется талломом. Различают два основных типа грибов: гифальный и дрожжевой.
Гифальные (плесневые грибы) образуют ветвящиеся тонкие нити (гифы). Гифы (от греч. hypha, паутина) представляют собой разветвленные микроскопические трубки диаметром 2-10 мкм, содержащие цитоплазму и органеллы. Совокупность гиф обозначают термином мицелий. Образование мицелия — отличительный признак истинных грибов (Eumycota). Гифы высших грибов содержат перегородки (септы), разделяющие их на отдельные клетки. Септы имеют отверстия, позволяющие цитоплазме и отдельным органеллам перетекать из одной клетки в другую. Гифы низших грибов не имеют перегородок и называются ценоцитными, или асептированными. Таким образом, плесневый гриб представляет собой ценоцит — обширную территорию цитоплазмы с множеством ядер, располагающуюся в скоплении трубок-гиф. Врастающая в субстрат часть тела гриба, абсорбирующая питательные вещества — это вегетативный мицелий, а растущая на поверхности субстрата часть — воздушный мицелий. Воздушный мицелий придает поверхности колоний плесневых грибов характерную шерстистую или пушистую фактуру. Нередко воздушный мицелий образуют специализированные гифы, несущие репродуктивные структуры (рис.26).

Рис. 26 Плесневые грибы рода Mucor
Дрожжевые грибы. Дрожжи и дрожжевые грибы представлены отдельными овальными клетками размером 3-10 мкм, морфологически сходными между собой. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий), состоящий из цепочек удлиненных клеток. Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называются дрожжевыми. Они размножаются только бесполым способом — почкованием или делением. На питательных средах дрожжи и дрожжевые грибы образуют блестящие, выпуклые колонии, сходные с колониями бактерий. Многие грибы обладают диморфизмом — способностью давать мицелиальный или дрожжеподобный рост в зависимости от различных факторов (например, условий культивирования).
У грибов выделяют половой и бесполый типы размножения. Бесполое размножение реализуется путем образования конидий и спорангиоспор, содержащих весь генетический материал, необходимый для возникновения и развития новой колонии. Конидии — неполовые репродуктивные структуры. Они могут быть одноклеточными (микроконидии) и многоклеточными (макроконидии). У некоторых грибов конидии формируются на концах конидионосных гиф — конидиофоров (бластоконидии), у других — по бокам и на концах обычных гиф (таллоконидии) путем их септирования и расчленения. Бластоспоры формируются путем отпочковывания от конидиофора, что характерно для дрожжей и дрожжевых грибов. Наиболее типичные таллоконидии — артроконидии и хламидоконидии. Артроконидии (артроспоры) образуются в результате фрагментации септированных гиф, распадающихся на отдельные клетки. Хламидоконидии (хламидоспоры) образуются внутри нитей мицелия в виде увеличенных клеток, окруженных толстой оболочкой. За счет плотной оболочки хламидоконидии приобретают устойчивость к различным воздействиям и сохраняются после гибели и распада мицелия (рис.27).

Рис. 27 Способы размножения грибов
Спорангиоспоры — споры, образующиеся вегетативным путем в специализированных мешках (спорангиях). Последние располагаются на концах особых гиф — спорангиофорах. К основным типам спорангиев относятся фиалиды (образования в форме бокала), и аннелиды (кольцевидные структуры).
Половым путем образуются аскоспоры, базидиоспоры и зигоспоры.
Среди грибов, имеющих медицинское значение, выделяют три типа (Phylum) или отдела, имеющие половой способ размножения (совершенные грибы): зигомицеты (Zygomycota), аскомицеты (Askomycota) и базидиомицеты (Basidiomycota). Кроме того, выделяют условный тип/группу грибов — дейтеромицеты (Deiteromycota), у которых имеется только бесполый способ размножения (несовершенные грибы).
14. Особенности строения и размножения актиномицет.
Актиномицеты — это микроорганизмы, занимающие промежуточное положение между бактериями и грибами. Длительное время актиномицеты считали грибами, однако изучение морфологии и биологических свойств позволило отнести их к бактериям семейства Actinomycetaceae отдела Firmicutes. Сходство с грибами определяется способностью к образованию на питательной среде субстратного и воздушного мицелия, на концах которого образуются споры; споры актиномицет являются способом размножения. Однако, в отличие от грибов, клеточная стенка актиномицет не содержит хитина или целлюлозы; они не способны к фотосинтезу, а образуемый ими мицелий достаточно примитивен. Они также резистентны к противогрибковым препаратам. С бактериями актиномицеты объединяет отсутствие четко выраженного ядра, наличие клеточной стенки, а также чувствительность к бактериофагам и антибиотикам.
Актиномицеты — ветвящиеся, нитевидные или палочковидные грамположительные бактерии. Свое название они получили в связи с образованием в пораженных тканях друз — колоний, состоящих из гранул плотно переплетенных нитей в виде лучей, отходящих от центра и заканчивающихся колбовидными утолщениями. Некислотоустойчивы. Факультативные анаэробы. Актиномицеты могут делиться путем фрагментации мицелия на клетки, похожие на палочковидные и колбовидные бактерии. Споры актиномицетов обычно нетермостойки.
Большинство актиномицет обитают на поверхности слизистых оболочек у млекопитающих; некоторые виды — почвенные сапрофиты. У человека актиномицеты колонизируют слизистые оболочки полости рта и ЖКТ. Способность вызывать специфические поражения не выражена и их рассматривают как условные патогены. Бактерии вызывают актиномикозы — хронические гнойные гранулематозные поражения различных органов.
Подавляющее большинство случаев актиномикозов у людей вызывает A.israelii, в редких случаях A.naeslundii, A.odontolyticus, A.bovis, A.viscosus.

Рис. 20 Актиномицеты, окрашенные по Граму
15. Особенности строения, движения, окраски спирохет.
Спирохеты — тонкие, длинные, извитые (спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной сгибательными изменениями клеток. Спирохеты имеют наружную мембрану клеточной стенки, окружающую протоплазматический цилиндр с цитоплазматической мембраной. Под наружной мембраной клеточной стенки (в периплазме) расположены периплазматические фибриллы, которые, как бы закручиваясь вокруг протоплазматического цилиндра спирохеты, придают ей винтообразную форму (первичные завитки спирохет). Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Другой конец фибрилл свободен. Число и расположение фибрилл варьирует у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение (рис.24). При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками.
Аксиальная
Внешняя
фибрилла
мембрана Протоплазматический
цилиндр



Внешняя
мембрана
Аксиальная
фибрилла
Периплазматическое
пространство
Пептидогликан Цитоплазматическая
мембрана





Рис. 24. Двигательный аппарат спирохет
Спирохеты плохо воспринимают красители из-за большого количества липидов в оболочке. Их окрашивают по методу Романовского-Гимзы или серебрением, а в живом виде исследуют с помощью фазово-контрастной или темнопольной микроскопии. Спирохеты представлены тремя родами, патогенными для человека: Treponema, Borrelia, Leptospira (рис.25).
Трепонемы (род Treponema, вид T.pallidum, возбудитель сифилиса) имеют вид тонких штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками. Вокруг протопласта трепонем расположены фибриллы. Окрашиваются в бледно-розовый цвет по методу Романовского-Гимзы.
Боррелии (род Borrelia, B.recurrentis, возбудитель возвратного тифа) более длинные, имеют по 3-8 крупных неравномерных завитков и 8-20 фибрилл. Окрашиваются в сине-фиолетовый цвет по методу Романовского-Гимзы.
Лептоспиры (род Leptospira, вид L.interrogans, возбудитель лептоспироза) имеют завитки неглубокие и частые — в виде закрученной веревки (завитки первого порядка). Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв "S" или "С"; имеют две осевые нити. Окрашиваются в красно-розовый цвет по методу Романовского-Гимзы.
Таблица 7. Дифференциальные признаки спирохет
Номенклатура |
Инфекционное заболевание |
Размеры, кол-во и характер завитков |
Метод Романовского-Гимзы (цвет) |
Род Borrelia Вид B.recurrentis |
Возвратный тиф |
3-8 крупных неравномерных завитков |
сине-фиолетовый |
Род Treponema Вид T.pallidum |
Сифилис |
8-12 равномерных мелких завитков |
бледно-розовый |
Род Leptospira Вид L.interrogans |
Лептоспироз |
Неглубокие частые завитки 1-го и завитки 2-го порядка, придающие бактерии форму букв "S" или "С" |
красно-розовый |

Рис. 25 Спирохеты, окрашенные по методу Романовского-Гимзе
16. Особенности строения, выявления, химический состав микоплазм.
Микоплазмы — мелкие полиморфные микроорганизмы (0,15-1,0 мкм), лишенные клеточной стенки и относящиеся к классу Mollicutes. Поверхностной оболочкой микоплазм является цитоплазматическая мембрана, но более прочная и эластичная, что связано с присутствием в ней холестерина. Большинство видов микоплазм нуждается в экзогенном холестерине или других стеринах для роста. Клетки микоплазм содержат нуклеоид (геном микоплазм является самым маленьким среди всех саморепродуцирующихся живых клеток), рибосомы, цитоплазму и цитоплазматическую мембрану. Иногда вокруг крупных морфологических элементов в электронном микроскопе виден мукозный слой, подобный капсуле. У некоторых микоплазм обнаружены микроворсинки и нитчатые или стеблеобразные выросты различной длины, которые принимают участие в скользящем движении клеток и адгезии. У микоплазм наблюдаются различные способы размножения: бинарное деление, фрагментация крупных тел и нитей с образованием мелких зерен, процесс, сходный с почкованием. Из-за отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют разнообразную полиморфную форму: кокковидную, нитевидную, колбовидную; похожи на L-формы (рис.23). На плотной питательной среде образуют колонии, напоминающие яичницу-глазунью: центральная непрозрачная часть, погруженная в среду, и более прозрачная периферия в виде круга.
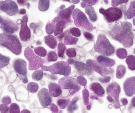
Рис. 23 Полиморфность микоплазм
Микоплазмы являются «мембранными паразитами», в основном поражают слизистые оболочки, где отсутствуют микробы-антагонисты. Они прочно прикрепляются к мембране клеток, усваивая из мембраны необходимые им питательные вещества, эпителиоциты при этом повреждаются, но не погибают. Микоплазмы изучают в нативных препаратах с использованием фазово-контрастной микроскопии, РИФ и генетических методов (ПЦР).
У человека микоплазмы вызывают микоплазмозы: M.pneumoniae (заболевания респираторного тракта), M.hominis, M.genitalium, U.urealyticum (инфекции урогенитального тракта).
17. Особенности строения, размножения риккетсий.
Риккетсии — мелкие, грамотрицательные полиморфные бактерии (0,3-2,0 мкм), облигатные внутриклеточные паразиты. Капсул и спор не образуют. Жизненный цикл риккетсий включает две стадии — вегетативную (внутриклеточную) и покоящуюся. Вегетативные формы активно размножаются бинарным делением в цитоплазме, а некоторые — в ядре инфицированных клеток, используя энергетические системы клетки-хозяина, поскольку не способны синтезировать кофермент НАД и др. метаболиты. Покоящаяся форма обладает повышенной резистентностью с утолщенной клеточной стенкой и уплотненной цитоплазмой.
Обитают в организме членистоногих (вшей, блох, клещей), которые являются их хозяевами или переносчиками. Форма и размер риккетсий могут меняться (клетки неправильной формы, нитевидные, кокковидные, бациллярные) в зависимости от условий роста. В мазках и тканях их окрашивают по Романовскому-Гимзе, по Здродовскому (рис.21) (риккетсии красного цвета, а инфицированные клетки — синего).
Таксономия риккетсий основывается на сравнении фенотипических, в том числе антигенных характеристик, клинико-эпидемиологических особенностях болезней, а также молекулярно-генетических данных, присущих отдельным представителям риккетсий. До недавнего времени семейство риккетсии включало роды Rickettsia, Orientia, Ehrlichia и Coxiella. По современной классификации (Е. П. Лукин, А. А. Воробьев, А. С. Быков, 2001), семейство Rickettsiaceae относится к классу Alphaproteobacteria (альфа-1 протеобактерии) и включает три рода: Rickettsia, Orientia, и Ehrlichia. Род Coxiella исключен из семейства Rickettsiaceae и отнесен к гамма-протеобактериям (близкое родство к легионеллам).
Патогенные для человека риккетсии вызывают риккетсиозы; среди них выделяют группы сыпных тифов (R.prowazekii, R.typhi) и пятнистых лихорадок (R.ricketsii, R.conorii), Ку-лихорадку (C.burnetii) и др.

Рис. 21 Риккетсии, окрашенные по методу Здродовского
18. Особенности строения и размножения хламидий.
Хламидии относятся к облигатным внутриклеточным кокковидным грамотрицательным (иногда грамвариабельным) бактериям. Они размножаются только в живых клетках. Вне клеток хламидии имеют сферическую форму (0,3 мкм), метаболически неактивны и называются элементарными тельцами. В клеточной стенке элементарных телец имеется главный белок наружной мембраны и белок, содержащий большое количество цистеина. Элементарные тельца попадают в эпителиальную клетку путем эндоцитоза с формированием внутриклеточной вакуоли. Внутри клеток они увеличиваются и превращаются в делящиеся ретикулярные тельца, образуя скопления в вакуолях (включения). Из ретикулярных телец образуются элементарные тельца, которые выходят из клеток путем экзоцитоза или лизиса клетки. Вышедшие из клетки элементарные тельца вступают в новый цикл, инфицируя другие клетки (рис.22).
Согласно последним данным семейство Chlamydiaceae включает два рода: собственно Chlamydia и новый род Chlamydophila; в последний входят виды C.pecorum, C.pneumonia и C.psittaci. Род Chlamydia включает, кроме C.trachomatis, два новых вида: C.muridarum и C.suis, выделенных соответственно от мышей, золотистых хомячков и от свиней.
Изучают хламидии в живом состоянии с использованием фазово-контрастной микроскопии и окрашивают по методу Романовского-Гимзы (элементарные тельца окрашиваются в розовый, ретикулярные — в сине-голубой цвет), а также в реакции иммунофлюоресценции (РИФ) и с помощью генетических методов (ПЦР).
У человека вызывают хламидиозы: C.trachomatis (возбудитель трахомы, урогенитальных инфекций), C.psittaci (орнитоз), C.pneumoniae (различные формы респираторных инфекций).

Рис. 22 Жизненный цикл хламидий
19. Основные представители патогенных простейших, их морфологические особенности, методы окраски.
Простейшие или протисты (Protozoa), состоят из единственной эукариотической клетки. Снаружи тело простейших покрывает ригидная мембрана — пелликула. К ней прилегает внешний более плотный и гомогенный слой цитоплазмы — эктоплазма. У некоторых видов пелликула может содержать опорные фибриллы и даже минеральный скелет. Набор органелл, расположенных в более жидкой эндоплазме, идентичен клеткам многоклеточных животных организмов; исключением может быть наличие у некоторых видов нескольких ядер. Многие простейшие способны активно передвигаться за счет псевдоподий (выростов цитоплазмы), жгутиков и ресничек. В неблагоприятных условиях простейшие могут образовывать цисты.
Простейшие разных классов могут вызывать различные инфекционные заболевания (табл. 9). В последнее десятилетие резко возросло значение простейших — латентных паразитов организма человека, которые активизируются при снижении иммунной реактивности (врожденные и приобретенные иммунодефициты). К ним относятся возбудители токсоплазмоза, пневмоцистоза и др. Трихомонада является одним из наиболее распространенных возбудителей инфекций урогенитального тракта. Для идентификации с помощью световой микроскопии используют окраску по Романовскому-Гимзы (цитоплазма окрашивается в синий, ядро — в красный цвет).
Таблица 9. Простейшие, имеющие медицинское значение
Таксоны |
Представители |
Нозологические формы |
Тип Sarcomastigophorae |
||
Подтип Sarcodina (Саркодовые) |
Entamoeba hystolyticum |
Амебиаз (амебная дизентерия) |
Подтип Mastigophora (Жгутиконосцы) |
Лейшмании Leishmania spp |
Лейшманиозы |
Трипносомы Trypanosoma brucei |
Африканский трипаносомоз |
|
Лямблии Giardia lamblia |
Лямблиоз (диарея, мальабсорбция) |
|
Трихомонады Trychomonas vaginalis |
Трихомоноз (вагиниты, уретриты, простатиты) |
|
Тип Apicomplexa |
||
Класс Sporozoa (Споровики) |
Плазмодии P.ovale, P.vivax, P.malariae |
Малярия |
Токсоплазмы Toxoplasma gondii |
Токсоплазмоз |
|
Тип Ciliophora |
||
Класс Kinetofragminophorea |
Балантидии Balantidium coli |
Балантидиазная дизентерия |
Тип Microspora |
||
Класс Microsporea |
Микроспоридии Enterocytozoon spp. |
Микроспоридиоз |