

778
.pdfтеля переносят на предметное стекло и тщательно расправляют препаровальными иглами, затем на срез стеклянной палочкой наносят каплю бальзама и сверху осторожно опускают покровное стекло. Бальзам должен равномерно распределиться между стеклами и пропитать срез. После этого препарат подсушивают, чтобы бальзам затвердел.
Стекла с парафиновыми срезами проводят через те же жидкости и также заключают срезы в бальзам, следя за тем, чтобы они были хорошо расправлены. Тщательно приготовленные препараты, заключенные в нейтральный бальзам и защищенные от солнечного света, могут без изменения храниться многими десятилетиями. Для наглядного ознакомления со значением окрашивания берем следующий препарат.
Изготовленные препараты можно изучать в световом микроскопе при увеличении в 10-2000 раз, их можно долго хранить.
Процесс изготовления препарата с использованием парафина длится 10 суток, а с использованием целлоидина - более месяца.
Контрольные вопросы и задания
1.Группы гистологических красителей.
2.Определение и содержание гистологической техники.
3.Этапы изготовления гистологического препарата.
4.Каковы цели фиксации мазка?
5.Методы микроскопирования гистологических препаратов.
6.Методы исследования в гистологии.
7.Сущность, техника и практическое значение окраски по Гра-
му.
8.Значение и сущность окраски по Романовскому — Гимза.
9.Микроскопическая техника. Правила микроскопирования.
10.Разрешающая способность светового и электронного микро-
скопа.
31
ЦИТОЛОГИЯ
Цитология – наука о клетке, об общих закономерностях, присущих клеточному уровню организации живой материи.
Клеточная теория в ее современном виде включает следующие положения:
1)Клетка – наименьшая единица живого.
2)Клетки тканей различных организмов имеют общие принципы строения: они состоят из ядра и цитоплазмы, содержащей органоиды и включения, и обладают основными свойствами «живого» – обменом веществ и энергии, способностью расти и дифференцироваться, отвечать на раздражения, самовоспроизводиться.
3)Размножение клеток происходит только путем деления исходной клетки.
4)В многоклеточных организмах любая клетка является частью целого организма, они специализированы по строению и функции и объединены в составе тканей, органов и систем. Ядро включает хроматин, ядрышко, кариоплазму (нуклеоплазму) и выполняет две группы функций: генетическую и регуляторную. Цитоплазма клетки включает в себя прозрачную бесструктурную гиалоплазму (цитозоль), органеллы – живые и активно работающие части клетки и включения, представляющие собой пассивный запас каких-либо веществ.
Органеллы – постоянные составные части цитоплазмы, выполняющие определенные функции (рис.2). Одни из органелл присутствуют в каждой клетке организма и поэтому называются общими, другие – лишь в клетках определенного типа и называются специальными. К общим органеллам относят митохондрии, цитоплазматическую (эндоплазматическую) сеть, пластинчатый комплекс (аппарат Гольджи), лизосомы, рибосомы, центросомы, пероксисомы, микротрубочки, микрофибриллы, а к специальным - миофибриллы, нейрофибриллы, тонофибриллы, реснички, жгутики, микроворсинки.
32
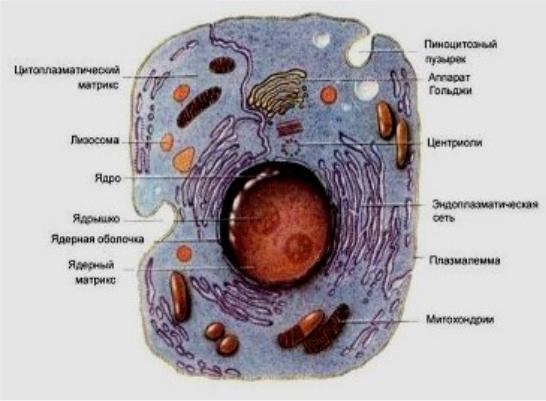
Рис.2. Схема строения животной клетки
Цитоплазма животной клетки отделена от окружающей среды и от соседних клеток – цитолеммой.
Цитолемма выполняет разграничительную функцию и регулирует движение ионов и молекул в клетку и из клетки, а также участвует в процессах фагоцитоза, пиноцитоза и экзоцитоза (рис.3). Она представляет собой элементарную биологическую мембрану, состоящую из бимолекулярного слоя липидов и белков - интегральных, полуинтегральных и периферических. Кроме того, с липидами и белками связаны молекулы углеводов, образуя с ними гликопротеиды и гликолипиды. Они формируют надмембранный комплекс – гликокаликс, в составе которого есть структуры, называемые рецепторами. Рецепторы способны специфически связывать определенные химические вещества: антигены, гормоны, медиаторы. С внутренней стороны мембраны располагается подмембранный комплекс, включающий в себя микрофиламенты, микрофибриллы и микротрубочки цитоскелета, а также сократительные белки.
33
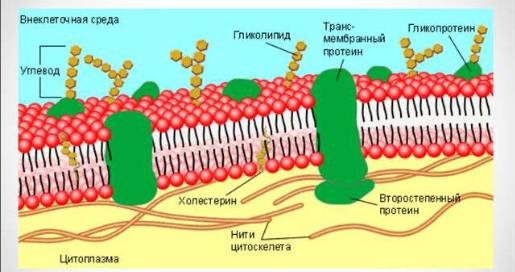
Рис.3. Схема строения цитолемм
Специализированными структурами цитолеммы являются различные типы межклеточных соединений, а также выросты цитоплазмы – микроворсинки, или более сложные по строению реснички и жгутики. Межклеточные соединения (контакты) подразделяют на замыкающие, адгезионные и коммуникационные (проводящие).
Кадгезионным контактам, механически скрепляющим клетки между собой, относятся десмосомы, полудесмосомы и опоясывающие десмосомы.
Ккоммуникационным контактам относятся щелевые контакты, через которые проходят низкомолекулярные вещества, регулирующие рост и развитие клеток, нексусы и синапсы, обеспечивающие передачу сигналов с одной клетки на другую.
Система биологических мембран клетки включает не только цитолемму и кариолемму (в составе которой две биологические мембраны с перинуклеарным пространством между ними), но и группу органелл мембранного строения: цитоплазматическую сеть, комплекс Гольджи, митохондрии, лизосомы и пероксисомы.
Обычно в нормально функционирующей зрелой клетке общее количество мембран постоянно, хотя структура и соотношение органоидов могут меняться.
34
Образование новых мембран в клетке идет с участием эндоплазматической сети двух видов: гладкой - образует липидную часть мембран, а гранулярная - их белковые компоненты. Эти процессы идут после деления клетки, при восстановительных процессах после повреждения, а также при гипертрофии клетки.
Органеллы (органоиды) общего значения - мембранного строения
Эндоплазматическая сеть (эндоплазматический рети-
кулум) представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих трехмерную мембранную сеть. В состав сети входят гранулярные и агранулярные участки, которые могут чередоваться.
Гранулярная эндоплазматическая сеть на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами (рис. 4).
Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазмати-
35
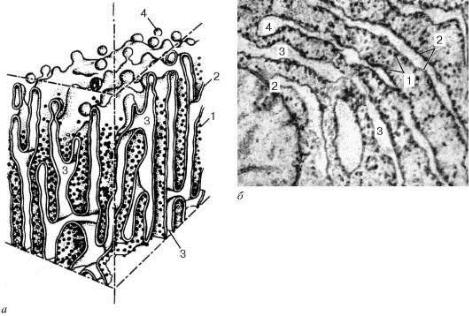
ческая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
Рис. 4. Строение гранулярной эндоплазматической сети:
а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки,
лишенные рибосом
36
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть также представлена мембранами, образующими мелкие вакуоли, трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50-100 нм. Гладкая эндоплазматическая сеть возникает и развивается на основе гранулярной эндоплазматической сети. В отдельных участках гранулярной эндоплазматической сети образуются новые липопротеидные мембранные участки, лишенные рибосом. Эти участки могут разрастаться, отщепляться от гранулярных мембран и функционировать как самостоятельная вакуолярная система.
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды, например, в эндокрин-
37
ных клетках коркового вещества надпочечников, в эпителиальных клетках извитых семенных канальцев.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при некоторых отравлениях в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь заполненные гладким эндоплазматическим ретикулумом.
Комплекс Гольджи (пластинчатый комплекс) был от-
крыт в 1898 г. К. Гольджи. Автор, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом. В дальнейшем его стали называть аппаратом, или комплексом Гольджи. Подобные структуры затем были описаны во всех клетках эукариот.
При рассмотрении в электронном микроскопе комплекс Гольджи представлен мембранными структурами, собранными вместе в небольших зонах (рис. 5).
Отдельная зона скопления этих мембран называет-
ся диктиосомой (стопкой Гольджи). Таких зон в клетке мо-
жет быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми находятся тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения - ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне ком-
38
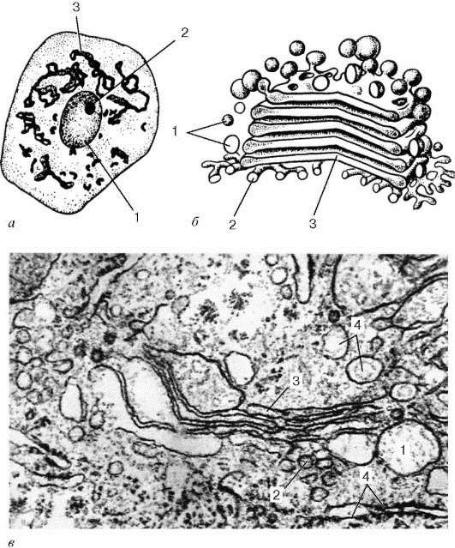
плекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются, главным образом, в его периферических участках. Иногда они отшнуровываются от ампулярных расширений на краях плоских цистерн. В зоне диктиосомы различают проксимальную и дистальную поверхности. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная поверхность обращена к ядру, в то время как дистальная - к поверхности клетки.
Рис. 5. Комплекс Гольджи:
а- нервная клетка спинного мозга, импрегнация серебром по методу Гольджи: 1 - ядро; 2 - ядрышко; 3 - комплекс Гольджи; б - схема
ультрамикроскопического строения (трехмерная реконструкция);
в- комплекс Гольджи на ультратонком срезе (печеночная клетка):
1- пузырьки; 2 - трубочки; 3 - уплощенные мешочки (цистерны); 4 - фрагменты гранулярной эндоплазматической сети
39
Вклетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к дистальной поверхности, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах.
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в эндоплазматической сети,
вих химической перестройке, созревании; в его цистернах происходят синтез полисахаридов, их комплексирование с белками, что приводит к образованию пептидогликанов. С помощью комплекса Гольджи осуществляется процесс выведения готовых секретов за пределы секреторной клетки.
Кроме того, комплекс Гольджи обеспечивает формирование лизосом. Мембраны комплекса образуются путем отщепления мелких вакуолей от гранулярного эндоплазматического ретикулума. Эти вакуоли поступают в проксимальный отдел комплекса Гольджи, где и сливаются с его мембранами. Следовательно, в комплекс Гольджи поступают новые порции мембран и продуктов, синтезированных в гранулярном эндоплазматическом ретикулуме. В мембранных цистернах комплекса Гольджи происходят вторичные изменения в структуре белков, синтезированных в гранулярном эндоплазматическом ретикулуме. Эти изменения (модификации) связаны с перестройкой олигосахаридных цепочек синтезированных гликопротеидов. Внутри полостей комплекса Гольджи с помощью различных ферментов по-разному модифицируются лизосомные белки и белки секретов: происходят последовательная замена и наращивание олигосахаридных цепочек. Модифицирующиеся белки переходят от цистерны проксимальной цис-поверхности в цистерны дистальной поверхности путем эстафетного переноса мелких вакуолей, содержащих транспортируемый белок.
Вцистернах дистальной поверхности происходит сортировка белков: на внутренних поверхностях мембран ци-
40