

семестр 1 / Бендик К.Ю. 4 група2 курс
.docxБендик К.Ю. 4 група 2 курс
Факультет ветеринарної медицини 211
Механізм м'язового скорочення
Передача збудження від рухових нейронів до м’язових волокон відбувається через медіатор ацетилхолін (АХ). Взаємодія АХ з холінорецегором кінцевої пластинки призводить до активації АХ-чутливих каналів і появи потенціалу кінцевої пластинки, який може досягати 60 мВ. При цьому область кінцевої пластинки стає джерелом дратівної струму для мембрани м'язового волокна, і на ділянках клітинної мембрани виникає потенціал дії (ПД), який поширюється в обидві сторони зі швидкістю приблизно 3-5 м/с при температурі 36 ° С.
Поширюючись по зовнішній мембрані, ПД надходить у м’язові волокна, де через бічну систему канальців транспортується до саркоплазматичного сальника, який служить сполучною ланкою між поверхневою мембраною та пристроєм скорочення м’язового волокна. Електрична стимуляція місця контакту призводить до активації ферментів, розташованих в місці контакту, та утворення інозітолтрифосфату. Інозітолтрифосфат активує кальцієві канали мембран термінальних цистерн, що призводить до виходу іонів Са 2+ з цистерн і підвищенню внутрішньоклітинної концентрації Са 2+ . Оскільки концентрація кальцію в саркоплазмі становить менше 10 ~ 7 моль/1 л, а в саркоплазматичній сітці-більше 10 ~ 4 моль/1 л, її іони починають виділятися у саркоплазму у великих кількостях. Вивільнений кальцій ініціює скорочення м’язів. Рівень кальцію досягається протягом 12-15 мілісекунд після надходження нервового імпульсу, чого достатньо для початку скорочення м’язів. Це прихований, латентний, час м'язового скорочення. У зв'язку з тим, що швидкість поширення ПД сарколемою більша від часу, необхідного для виділення Са2+ із саркоплазматичного ретикула, усі фібрили ділянки м'яза, що іннервуються одним нервом, скорочуються одночасно. Сукупність процесів, що призводять до збільшення внутрішньоклітинної концентрації Ca 2+, є суттю третьої фази скорочення м’язів. Тому на першому етапі електричний сигнал ПД перетворюється на хімічний сигнал - збільшення внутрішньоклітинної концентрації Са 2+.
При підвищенні внутрішньоклітинної концентрації іонів Са 2+ тропоміозин зміщується в жолобок між нитками актину, при цьому на актинових нитках відкриваються ділянки, з якими можуть взаємодіяти поперечні містки міозину. Ця зміна тропоміозину обумовлена зміною конформації молекули білка тропоніну під час зв'язування Са 2+. Отже, участь іонів Са 2+ в механізмі взаємодії актину і міозину опосередковано через тропонін і тропоміозин.
Наступним етапом електромеханічного сполучення є приєднання головки поперечного містка до актинових філаментів, до першого з декількох послідовно розташованих стабільних центрів. Головка міозину обертається навколо своєї осі, оскільки має кілька центрів активності, які, у свою чергу, взаємодіють з відповідними центрами на нитках актину. Обертання головки приводить до збільшення пружної еластичної тяги шийки поперечного містка і збільшення напруги.
У кожен конкретний момент в процесі розвитку скорочення одна частина головок поперечних містків знаходиться в з'єднанні з актиновими філаментами, інша вільна. Між ними та актиновими нитками існує ряд взаємодій. Це забезпечує плавність процесу скорочення. На четвертому і п'ятому етапах відбувається механічне перетворення.
Послідовна реакція з'єднання і роз'єднання головок поперечних містків з пктиновими філаментами призводить до ковзання тонких і товстих ниток відносно один одного і зменшення розмірів саркомера і загальної довжини м'яза, що є шостим етапом. Сукупність описаних процесів становить сутність теорії ковзання ниток.
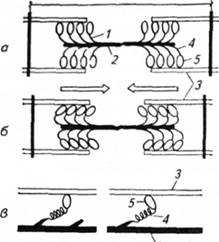
Схема діяльності поперечних містків (1) і міозинових протофібрил (2): а - поперечні містки в стані розслаблення м'язового волокна; 6 - під час скорочення (стрілками зазначено напрямок руху актинових протофібрил (і) двох половин саркомера); в - модель розвитку напруження в поперечних містках під час їхнього скорочення (ліворуч - у стані розслаблення, праворуч - під час скорочення м'язового волокна). 4 - шийка поперечного містка; 5 - головка поперечного містка
Після цього утворений фосфор та аденозиндіфосфат (АДФ) видаляються, а на їх місце додаються нові молекули АТФ, що призводить до розриву зв’язку між міозином та активним центром актину.При скороченні м'яза:
а) актинові й міозинові нитки практично не коротшають;
б) взаємодія актину з міозином призводить до взаємного входження ниток у проміжки між ними;
в) дві прилеглі 7-мембрани зближаються одна з одною, і при максимально сильному скороченні відстань між ними може зменшитися майже вдвічі;
г) при зменшенні довжини м'яза саркомер розширюється, позаяк укладена всередині саркомера саркоплазма не стискається;
ґ) подібні процеси одночасно перебігають у всіх саркомерах м'язового волокна, тому обидва кінці м'яза підтягуються до центру.
https://pidru4niki.com/1228112859765/meditsina/skeletni_myazi
https://studfile.net/preview/4513054/
https://elearning.sumdu.edu.ua/free_content/lectured:b40514a462ccfa8803aa0b3da100825c3ef6d739/20150923093602/115382/index.html
https://stud.com.ua/76664/meditsina/mehanizm_myazovogo_skorochennya
http://www.um.co.ua/11/11-2/11-28420.html
https://yogagorod.ru/uk/tipy-myshechnyh-sokrashchenii-kachaemsya-pravilno-izometricheskoe/
https://theroyalfamily.ru/uk/steny/sistema-organizma-kontroliruyushchaya-sokrashchenie-skeletnyh-myshc/