

Физиология и биофизика возбудимых тканей. 2
Регуляция физиологических функций. 42
Физиология эндокринной системы. 52
Физиология центральной нервной системы. 55
Физиология сердечно-сосудистой системы. 87
Физиология дыхания. 147
Физиология пищеварительной системы. 177
Физиология обмена веществ и терморегуляция. 209
Физиология выделения. 234
Физиология крови. 244
Физиология сенсорных систем. 271
Физиология высшей нервной деятельности. 306
Меня зовут Яковенко Никита Сергеевич, я написал этот учебник в 2020-м году для студентов ПИМУ. Сделал я это потому, что любой желающий студент имеет право на достойное образование, а нормальную физиологию важно знать и понимать очень хорошо всем будущим врачам, в связи с этим я постарался сделать учебник максимально полезным для вас. Волчара – это моя подруга Таня Волкова (девичья фамилия), она очень классная, и во многом учебник изначально был сделан для неё, поэтому она заслужила упоминания.
Данный учебник составлен по программе лечебного факультета, но большая часть программ других факультетов здесь тоже имеется (они не сильно-то отличаются). При составлении учебника я постарался учесть всё, что спрашивают преподаватели на парах и на экзамене. При пользовании учебником ориентируйтесь на вопросы рабочей тетради, которую вы получаете на кафедре (там есть раздел «вопросы для самоподготовки» - по ним и спрашивают). Лишнего материала здесь нет, а главы расположены в том порядке, в котором они идут (ну или шли) в учебной программе, поэтому рекомендую читать всё подряд. Особенности учебника:
Материал подобран непосредственно под программу этого вуза, т.е. есть вся нужная информация и нет лишней. Правда надо учесть, что с годами программа несколько меняется, поэтому некоторых мелочей вы можете не найти, однако около 90-95% информации здесь точно есть, так как на данном курсе вы проходите основы нормальной физиологии, а информация об основах с течением времени меняется не сильно, редко.
Информация здесь объясняется так, чтобы она была действительно понятна всем, кто этого желает.
Касательно того, важности предмета. На младших курсах, и вообще в медицинском, есть четыре главных предмета: биохимия, физиология, патологическая физиология и патологическая анатомия. Данные предметы создают общее представление о работе организма и о его патологиях. Все клинические дисциплины базируются на этих предметах, поэтому они вам будут крайне полезны (изучать частные патологии, не усвоив общих, довольно проблематично, как и изучение общей патологии без предварительного усвоения общего нормального функционирования организма). В данных предметах, разумеется, главным является понимание – эти предметы на логическое мышление. Если вы хорошо их освоите, то они послужат очень хорошей основой для структурного и клинического мышления, которое врачу очень необходимо и развивается непросто.
Касательно того, как учить. Этот предмет в списке приоритетов желательно поставить повыше (он того стоит), учить его к каждой паре (навёрстывать тяжело), учить не в последний день, а хотя бы в предпоследний (стоит подстраховаться, ибо вы можете не успеть, так как предмет непростой).
Физиология и биофизика возбудимых тканей.
Введение.
Перед тем, как приступать к конкретной физиологии, нужно обговорить некоторые азы (темы вводного занятия).
Физиология изучает процессы жизнедеятельности и механизмы их регулирования в различных биологических системах (от клетки до организма). Предметами изучения физиологии являются функции живого организма, взаимосвязь этих функций, их регуляцию, приспособление организмов к внешней среде и становление их в филогенезе и онтогенезе. Физиологию разделяют на общую (функции клеток), частную (функции тканей) и прикладную (функции целого организма), второй подход в классификации: нормальная, патологическая, клиническая. Мы будем разбирать нормальную.
Основные понятия, которые надо усвоить перед изучением раздела возбудимых тканей: раздражитель, раздражение, раздражимость, возбуждение, возбудимость.
Раздражитель – фактор среды, действующий на живую систему, вызывая в ней ответ в виде изменения обмена веществ. Раздражение – процесс изменения обмена веществ в живой системе в ответ на действие раздражителя. Раздражимость – способность биологической системы отвечать изменением обмена веществ на действие раздражителя. В общем это всё об одном и том же, только надо чётко отличать между собой фактор, процесс и способность. Раздражимость – в каком-то смысле основа живой природы, это свойство позволяет отдельным объектам получать некоторую независимость и обособленность от стихийных процессов, именно этим живая природа отличает от неживой (есть и другие свойства, но это ключевое).
Однако в некоторых тканях раздражитель может вызывать необычную реакцию – возбуждение. Возбуждение – процесс быстрого колебания мембранного потенциала покоя МПП с изменением знака заряда мембраны в ответ на действие раздражителя. Возбудимость – способность биосистемы быстро колебать МПП с изменением знака заряда мембраны в ответ на действие раздражителя. Подробнее эти вопросы разберём чуть позже, но суть в том, что это электрические процессы, то есть в тканях в прямом смысле возникает ток, который может передаваться по тканям, т.е. мы похожи на машины, компьютеры в каком-то смысле. Возбудимыми являются только нервная, мышечная и секреторная (секреторные клетки эпителиальной) ткани.
В данном разделе мы будем разбирать электрические явления в клетках и тканях, за формирование этих явлений в клетке отвечает цитоплазматическая мембрана, поэтому перед тем, как разбирать электрические явления, надо изучить строение и функции цитоплазматической мембраны.
Цитоплазматическая мембрана состоит из двух компонентов – белков и липидов. Все липиды в мембране амфифильные, то есть имеют полярную (гидрофильную) и неполярную (гидрофобную) части, полярные части обращены к цитоплазме и тканевой жидкости, а неполярные части обращены друг к другу – так формируется двойной слой липидов. Основные липиды: глицерофосфолипиды, сфингофосфолипиды, гликосфинголипиды и холестерол. Липиды образуют основу мембраны (мембраны без липидов не бывает), позволяют белкам закрепляться на мембране и позволяют им принять нативную структуру, формируют барьер (далеко не все вещества способны пройти через билипидный слой), участвуют в передаче сигналов, выполняют защитную функцию (формируют гликокаликс), позволяют мембране быть одновременно текучей (длинные радикалы и ненасыщенные жирные кислоты) и твёрдой (холестерол и насыщенные жирные кислоты). липиды по большей части отвечают за структуру мембраны, а белки – за функции. Белки бывают интегральными (проходят мембрану насквозь), полуинтегральными (частично погружены в мембрану) и поверхностными (примыкают к мембране, прикрепляясь/заякореваясь разными способами: прикрепляются к гликосфинголипидами ковалентно, имеют свою жирную кислоту, встраивающуюся в мембрану, прикрепляются по заряду и тд). Функции белков: транспортная, регуляторная, ферментативная, электрогенная и тд. Общая схема строения цитоплазматической мембраны см ниже.

Гораздо подробнее строение, функционирование и метаболизм мембран вы будите разбирать на биохимии, а для курса нормальной физиологии достаточно запомнить модель общего строения мембраны, её компоненты и функции (барьерная, транспортная, регуляторная, электрогенная, межклеточные взаимодействия). Однако из темы биологических мембран для курса нормальной физиологии необходимо подробно разобрать вопрос транспорта веществ (это пригодится в этой и других темах). Существует 4 вида транспорта: простая диффузия, облегчённая диффузия, активный транспорт и цитоз (везикулярный транспорт).
Простая диффузия. Все неполярные вещества (стероиды, непрямой билирубин, кислород и тд) и мелкие полярные молекулы (этиловый спирт, углекислый газ, аммиак) способны проходить через билипидный слой без помощи всяких переносчиков, поэтому это простая диффузия (диффузия – взаимопроникновение веществ, молекулы одного вещества проходят спокойно между молекулами другого). Данный вид транспорта считается пассивным, потому что вещество движется из области большей концентрации в область меньшей (движущая сила – градиент/разница концентраций), а энергия для этого не затрачивается (поэтому и пассивный).
Облегчённая диффузия. В отличии от неполярных и мелких полярных веществ, крупные полярные и все заряженные вещества не способны диффундировать через билипидный слой, поэтому, чтобы пройти через клетку, им нужны специальные белки-переносчики. Кстати в этом случае вещество так же движется под градиентом концентраций, поэтому не надо тратить энергию – такой транспорт считается пассивным. Существует два основных вида переносчиков для облегчённой диффузии: транслоказы и каналы. Транслоказы, несмотря на окончание «аза», ферментами не являются; эти белки проходят мембрану насквозь, имеют центр связывания с конкретной молекулой (высокая специфичность), если эта молекула присоединится к центру, конформация белка серьёзно изменится, белок провернётся, перенося молекулу на другую сторону мембраны, молекула отсоединится, а переносчик примет изначальную конформацию, вернув активный центр на прежнее место и будет готов принять новую молекулу. Транслоказы могут переносить одну молекулу (пассивный унипорт), две разные молекулы в одну сторону (пассивный симпорт), две разные молекулы в противоположные стороны (пассивный антипорт). А как работают каналы? Эти белки тоже прошивают мембрану насквозь и имеют канал, т.е. буквально дыру в мембране (у транслоказ такого нет), вот только внутри канала выстилка из аминокислот бывает разная, поэтому каналы отличаются друг от друга по свойствам. Какими же бывают каналы? По пропускной способности: селективные (пропускают строго определённые вещества) и неселективные (пропускают всё, что проходит по диаметру). По воротам: безворотные (открыты 24/7) и воротные (они закрыты, но могут открыться, если их включить, т.е. это регулируемые каналы, могут иметь 1-2 ворот). Воротные каналы по способу рецепции делятся на механозависимые (рецептор канала воспринимает механическое воздействие, например, растяжение клетки, и включает канал), потенциал зависимые (рецептор канала реагирует на изменение заряда мембраны клетки, включая канал) и лиганд зависимые (рецептор реагирует на присоединение к нему химического вещества, например, нейромедиатора). Также воротные каналы классифицируются по способу связи рецептора канала и самого канала: ионотропные (рецептор приделан к самому каналу – реакция мгновенная) и метаботропные (рецептор канала находится на удалении от канала и активирует его через ряд вторичных посредников, каналы с такими рецепторами реагируют с задержкой, подробнее в курсе биохимии).
Активный транспорт. Некоторые вещества в клетке есть необходимость переносить против градиента концентрации, но сами просто так молекулы этого делать не станут (в гору не пойдут), поэтому нам приходится тратить энергию (гидролиз АТФ), чтобы переносить вещества против градиента их концентрации. Выделяют первично и вторично активный транспорт. В чём разница? При первично активном транспорте переносчик непосредственно тратит АТФ, например, натрий-калиевая-АТФаза тратит одну АТФ, чтобы перенести из клетки в тканевую жидкость 3 молекулы натрия, а из тканевой жидкости в клетку 2 молекулы калия, при этом работает она похоже на транслоказу, но с затратой АТФ. Такие переносчики по аналогии называют активными уни-, сим- и антипортами. Подробнее работа данного переносчика разбирается в курсе биохимии. А как работает вторично активный транспорт? Разберём на примере переносчика глюкозы в почечном канальце, который переносит глюкозу из просвета канальца нефрона в эпителиальную клетку этого нефрона против градиента концентрации. Вне клетки есть натрий, который по градиенту (вне клетки натрия гораздо больше, чем в клетке) хочет зайти в клетку, переносчик глюкозы разрешает ему это сделать только при условии, что натрий прихватит с собой глюкозу и протащит её против градиента её концентрации. Казалось бы, перенесли глюкозу против градиента концентрации, не потратив АТФ, но это не совсем так – для поддержания градиента концентраций натрия (именно он позволяет работать переносчику глюкозы) должна работать натрий-калиевая-АТФаза, которая постоянно выкачивает натрий из клетки, тратя АТФ. Таким образом, при вторично активном транспорте сам переносчик не тратит энергию АТФ, однако для работы этого переносчика АТФ тратят другие переносчики (опосредованная трата АТФ = вторичный транспорт), а при первичном транспорте переносчик непосредственно тратит АТФ.
Цитоз, он же везикулярный транспорт. Выделяют эндоцитоз (из тканевой жидкости в клетку) и экзоцитоз (из клетки в тканевую жидкость), эндоцитоз подразделяется на пиноцитоз (захват чего-то более-менее жидкого, растворимого, не очень большого) и фагоцитоз (захват крупных частиц, даже целых клеток). При эндоцитозе происходит впячивание мембраны, а затем отшнуровка, в результате часть тканевой жидкости с её содержимым оказывается в клетке в спец упаковке. Такой транспорт используется для объектов, которые нельзя транспортировать иначе, как правило, это все крупные молекулы и объекты. Но клетка не на любом участке мембране способна к таким действиям – у клетки на мембране есть специальные участки/площадки, под которыми есть спец белки, позволяющие мембране впячиваться и отшнуровываться, формирую везикулу/пузырёк, а самое главное – в этих участках есть рецепторы, запускающие эндоцитоз, распознавая крупные молекулы.
Благодаря тому, что билипидный слой непроницаем для большинства молекул, и тому, что транспорт большинства молекул осуществляется специфическими переносчиками, мембрана обладает избирательной проницаемостью, т.е. пропускает через себя только определённые молекулы в определённом количестве в определённых условиях по мере своих потребностей. Это очень важное свойство мембраны, позволяющее обмениваться клетке с внешней средой или другими клетками веществами, энергией и информацией (главные свойства живого).
Биотоки.
Теперь перейдём к электрическим явлениям в клетках и тканях нашего организма и начнём с открытия биотоков. В наше время электрический ток – неотъемлемая часть жизни, а несколько сотен лет назад ток был чем-то чуть ли не сверх естественным, и естественно людям хотелось думать, что такой чудесной вещи имеет место присутствовать и в человеке (вспомните, как доктор Франкинштейн подарил жизнь своему чудовищу), поэтому Гальвани (его мотивы были не столь примитивны, как я сейчас описал) решил доказать существование электрического тока в живых организмах (биотоки). Для этого он приготавливал спинальный препарат из лягушек (отрезал всё, что выше подмышек, снимал кожу с нижних лапок), вешал его за спину на медный крючок, который стоял на цинковой пластине. Когда лапка касалась пластины, лапка одёргивалась (ток вызывает сокращение мышц). Казалось бы, доказал, но Ватт выяснил, что источником тока послужила не лягушка, а так называемый гальванический элемент (соединение меди с цинком). Тогда Гальвани решил сделать другой эксперимент: он приготовил нервно-мышечный препарат (отпрепарированная мышца и подходящий к ней нерв), он накинул нерв на мышцу, после чего та начала сокращаться. В чём дело? Дело в том, что мембранный потенциал покоя у мышцы отрицательный (позже объясню, что это такое, а пока обратите внимание на «отрицательный потенциал»), но Гальвани так отпрепарировал мышцу, что часть её поверхности была целая (способна поддерживать физиологически нормальный отрицательный потенциал), а другая часть была деформирована (неспособна поддерживать потенциал отрицательным, поэтому потенциал равен нулю) – выходит, что на мышце имеются места с разными потенциалами (разными, потому что -70 мВ, например, и 0 мВ – разные потенциалы), а электрический ток по определению – разность потенциалов. Значит, частично повреждённая мышца генерирует электрический ток, который стимулирует наложенный на неё нерв, а тот, в свою очередь, вызывает сокращение мышцы. Так Гальвани своим вторым опытом доказал, что в живых организмах есть электрические токи (биотоки). Описанное явление было названо током покоя. Ток покоя – ток, возникающий между повреждённым и неповреждённым участками мембраны (цитоплазматическая мембрана клеток имеется ввиду) в покое (именно в покое, не в электротон, не в локальный ответ, не в потенциал действия, а именно в покое!). Однако на этом история не заканчивается, учёный Маттеучи взял два нервно-мышечных препарата, на первую мышцу положил нерв от второй мышцы, а на нерв от первой мышцы стимулятором подал ток, первый нерв возбудился, вызвал возбуждение и сокращение первой мышцы, а возбуждённая мышца простимулировала лежащий на ней нерв от второй мышцы, тот возбудил и заставил сокращаться вторую мышцу. Что произошло? В этот раз мышцы были отпрепарированы осторожно – все мембраны были целыми, дабы не вызвать ток покоя, поэтому первая мышца не стимулировала нерв от второй мышцы (между ними не было разницы потенциалов, а, следовательно, и тока), но возбуждённая первая мышца стимулировала изначально невозбуждённый нерв от второй мышцы, т.е. возбуждённая мышца имеет иной потенциал, нежели покоящаяся мышца и нерв – появилась разница потенциалов и, соответственно, электрический ток. Такой ток был назван током действия. Ток действия – ток, между возбуждённым и невозбуждённым участками мембраны (в покое и возбуждение мембрана имеет разные потенциалы, а разность потенциалов и есть электрический ток).

Мы уже не раз упоминали мембранный потенциал покоя (МПП – запоминайте аббревиатуры). Пришло время разобраться в том, что он из себя представляет. МПП – разность потенциалов между внутренней и внешней стороной мембраны. Только, пожалуйста, не путайте заряд мембраны и потенциал, в покое заряд мембраны положительный снаружи и отрицательный внутри, а потенциал отрицательный (заряд с внешней стороны и потенциал обратны по знаку, а заряд с внутренней стороны и потенциал равны по знаку). Потенциал – динамический показатель, характеризующий биотоки. За счёт чего формируется МПП? Во-первых, за счёт существования градиентов концентрации ионов натрия, калия, хлора. Во-вторых, за счёт высокой избирательной проницаемости мембраны для калия. В-третьих, за счёт функционирования натрий-калиевой-АТФазы.

Натрий и хлорид – осмотически активные ионы, т.е. они притягивают воду, клетка всеми силами пытается от них избавиться, поэтому их концентрация вне клетки явно выше, чем внутри клетки. Если бы клетка наполнилась ионами натрия и хлора, то она переполнилась бы водой и лопнула в прямом смысле. Зато ионы калия не являются осмотически активными, поэтому клетка старается их накапливать (они не грозят ей переполнением водой), поэтому концентрация калия в клетке гораздо выше, чем вне её. Отсюда мы имеем градиенты концентрация для трёх ионов, а градиент концентрации – движущая сила для облегчённой диффузии.
Что касается высокой избирательной проницаемости мембраны для калия. В цитоплазматической мембране есть безворотные каналы для калия, которые позволяют калию постоянно выходить из клетки без всяких препятствий, а натрий через этот канал может пролезть в клетку, но совсем чуть-чуть, незначительно. Есть, конечно, специальный канал для натрия и дополнительный канал для калия, но они с воротами и в покое закрыты, а мы обсуждаем мембранный потенциал именно покоя. А с натрий-калиевой –АТФазой вы уже знакомы.
Так что нам дают три этих фактора? Ну смотрите. За счёт высокого градиента (калий выходит по градиенту концентрации, так как вне клетки его меньше, но против электрического градиента, ибо он положительно заряжен, а выходит на положительно заряженную сторону мембраны, но это не сильно ему мешает выходить) калий выходит из клетки – из клетки убегают «плюсы». Затем с помощью натрий-калиевой-АТФазы калий возвращается в клетку, но в клетку входит два калия, а взамен выходит три натрия – два плюса вошло, а три вышло, т.е. клетка снова теряет «плюсы». В покоящемся состоянии клетка постоянно теряет «плюсы», поэтому её мембранный потенциал покоя отрицателен (для нервной ткани он составляет -70 мВ, для пеймейкеров -60 мВ, для типичных кардиомиоцитов -90 мВ). По одному из определений электрический ток – это направленное движение заряженных частиц. МПП – направленное (прочь из клетки) движение заряженных частиц (ионов, в первую очередь калия).

Мембранный потенциал покоя присущ всем клеткам и отличается лишь числовым значением. Для замера МПП вам потребуется микровольтметр и микроэлектроды. Один электрод помещается на поверхность мембраны, а второй проникает под мембрану клетки, а МПП – это разность потенциалов между внутренней и наружной стороной мембраны, поэтому мы размещаем электроды именно так. Мы пользуемся микровольтметром, ибо регистрируемый потенциал крайне мал, а в микровольтметре есть усилитель, который позволяет сделать замеряемый потенциал более ощутимым для прибора.
Теперь ещё одна не менее важная тема – потенциал действия. Потенциал действия – быстрое колебание МПП с изменением знака заряда мембраны в ответ на действие раздражителя. За формирование ПД будут отвечать двухворотные каналы для натрия, одноворотной канал для калия, натрий-калиевая-АТФаза. Как это всё происходит? В мембране есть потенциал чувствительные каналы для натрия, причём с разными настройками. Часть из них открывается при изменении потенциала на 1 мВ, часть при изменении на 2 мВ, и тд. Когда раздражитель действует на мембрану клетки, он уменьшает заряд мембраны, открывая постепенно каналы для натрия – график поднимается вверх (имеется ввиду, что уменьшается модуль заряда). Этот процесс – постепенное открытие разнонастроенных каналов для натрия, уменьшение заряда мембраны и как следствие изменение потенциала – называется медленной деполяризацией. Когда открываются каналы для натрия, тот входит в клетку по электро-химическому градиенту. Заметьте: когда мы обсуждали МПП, клетка теряла «плюсы», поэтому МПП был отрицательным, а во время медленной деполяризации «плюсы», т.е. натрий, входят в клетку, поэтому потенциал изменяется, становится менее отрицательным. Потенциал в покое отмечают на графике как Е-нулевое. Если раздражитель будет достаточным по ряду параметров, то потенциал дойдёт до отметки Е-критическое, это такое значение потенциала, при котором открываются все оставшиеся натриевые каналы и начинается быстрая деполяризация. В чём разница? В медленную деполяризацию по очереди открывалось небольшое количество натриевых каналов (в результате открылось, например, 40% каналов), натрий обильно поступал в клетку, заставляя потенциал изменяться в сторону более положительного, но в быструю деполяризацию открываются все оставшиеся 60% по принципу «всё или ничего» (абстрактно взятое число, не запоминайте – оно просто для понимания) одновременно, поэтому потенциал очень быстро изменяется в положительную сторону и даже изменяет знак (потенциал становится положительным, поэтому наружный заряд мембраны становится отрицательным – вот вам и перезарядка мембраны, о которой говорилось в определении ПД) – всё это происходит за счёт лавинообразного поступления натрия в клетку, он так повлиял на потенциал. Но у натриевых каналов есть особенность – они двухворотные, когда они включаются, у них открываются быстрые ворота (медленные изначально были открыты), спустя некоторое время захлопываются медленные ворота, тогда каналы считаются инактивированными (период инактивных ворот длиннее, чем период открытых ворот, т.е. допустим, канал был открыт 0,2 мс, а инактивированным после этого он будет 0,3 мс), инактивированный канал невозможно включить – приходится ждать, пока он станет снова просто закрытым; спустя некоторое время закрываются быстрые ворота, а медленные открываются – канал можно считать закрытым и готовым к работе. То есть усвойте: натривые каналы с двумя воротами бывают в трёх положениях: закрытые (быстрые ворота закрыты, медленные открыты, такой канал можно в любой момент открыть), открытые (обои ворота открыты), инактивированные (быстрые открыты, медленные закрыты, такой канал нереально открыть).
Но помимо всего этого, в начале медленной деполяризации открывались одноворотные каналы для калия, тот начинал выходить из клетки дополнительно, но во время быстрой и медленной деполяризации натрий входил в клетку куда больше, чем калий выходил, поэтому потенциал изменялся в положительную сторону – в клетку «плюсов» поступало больше, чем уходило. Но вот быстрая деполяризация закончилась, все натриевые каналы выключены, натрий резко перестал поступать в клетку, зато калий уже успел набрать обороты и обильно выходит из клетки через одноворотные каналы, поэтому потенциал резко изменяется в отрицательную сторону. Период изменения потенциала от пика до Е-критического называется быстрой реполяризацией, период от Е-критического до Е-нулевого называется следовой деполяризацией. Однако ток калия через одноворотные каналы бывает так силён, что порою вызывает следовую гиперполяризацию – график опускается ниже Е-нулевого, но потом возвращается в норму, потому что со временем все воротные каналы для калия закрываются. Что касается натриевых каналов, важно заметить, что во время быстрой реполяризации, пока потенциал положительный, все они инактивированы, т.е. их нереально включить, а вот когда потенциал становится отрицательным, часть натриевых каналов из инактивированных переходят в состояние закрытых (их можно активировать) и чем ближе быстрая реполяризация спускается к Е-критическому, тем больше натриевых каналов становятся закрытыми и готовыми к работе, в точке Е-критическое все натриевые каналы закрыты и готовы к работе. Натрий-калиевая-АТФаза работает постоянно, чтобы поддерживать градиенты концентраций натрия и калия (ПД запустил натрий в клетку, а АТФаза выкачивает, ПД выпустил калий из клетки, а АТФаза загоняет его обратно; если бы АТФаза перестала работать, после нескольких ПД концентрации ионов по обе стороны мембраны уравнялись, пропал бы градиент и ионы перестали бы бегать по каналам, вызывая колебание МПП). Вернёмся к определению ПД, это быстрое колебание МПП с изменением знака заряда мембраны – оно и видно, потенциал быстро колеблется в течение милисекунд, а то и быстрее, а мембрана на время меняет знак, потому что потенциал тоже на время меняет знак (на короткое время потенциал становится положительным, а наружный заряд мембраны отрицательным). Вот как раз перезарядка мембраны является визитной карточкой ПД (он же клеточный ответ, он же возбуждение). Наши нейроны передают по волокнам возбуждение в форме двоичного кода: отсутствие возбуждения (мембрана положительная) это 0, наличие возбуждения (мембрана отрицательная) это 1 – прям как у компьютеров.
Прошу заметить, что медленная деполяризация не входит в состав ПД либо считается нулевой фазой. Почему так? Потому что раздражитель может быть слабым или действовать недостаточно долго или ещё что, в результате этого медленная деполяризация возникнет, но не достигнет Е-критического и угаснет, тогда никакого ПД не будет. А вот остальные 4 фазы входят в состав ПД: быстрая деполяризация, быстрая реполяризация, следовая деполяризация, следовая гиперполяризация. Это общая схема, есть частные случаи, для которых будут совсем другие графики, в которых будут отсутствовать одни фазы и появятся другие, другие цифры, но это мы изучим значительно позже.

При построении графика обращайте внимание на единицы измерения (потенциал измеряется в миливольтах, а время в милисекундах) и на числовые значения! Также важно, чтобы линия быстрой деполяризации была круче, чем линия быстрой реполяризации, так как деполяризация быстрее, чем реполяризация. Получается, что каждая последующая фаза должна быть плавнее предыдущей. Также не забывайте на графике обозначать стимул/раздражитель, который запускает медленную деполяризацию. Это всё важные требования для того, чтобы ваш график был достаточно правдоподобным, в физиологии графики это святое.
В целом мы обсудили ПД и его природу (работа ионных каналов) по ходу всех фаз. Но для более полного понимания и закрепления материала предлагаю частные схемы (учить не надо – они для понимания).
Медленная деполяризация.

Я схематично изобразил мембрану с её каналами. В мембране всегда есть безворотные каналы для калия и натрий-калиевая-АТФаза для поддержания МПП, поэтому я их нарисовал по краям мембраны. За все фазы ПД по сути отвечают воротные каналы для калия (у них одни ворота на один канал, в покое закрыты) и воротные каналы для натрия (по двое ворот на канал, в покое быстрые ворота закрыты, медленные открыты). В покое все воротные каналы закрыты. Что происходит в медленную деполяризацию? Начинают открываться каналы для натрия постепенно (на рисунке открылись два канала), но вместе с тем начинают открываться каналы для калия (один канал на рисунке), однако ток натрия в клетку выше, чем ток калия из клетки, т.е. клетка наполняется «плюсами», поэтому мембранный потенциал изменяется в сторону положительного (график идёт вверх), пока потенциал не достигнет уровня Е-критического.
Быстрая деполяризация.

Как только мембранный потенциал достигает Е-критического, все оставшиеся натриевые каналы открываются (на рисунке их 4), в итоге, теперь вообще все натриевые каналы открыты (принцип «всё или ничего»). Воротные каналы для калия тоже открываются все, но не одновременно (хотя одновременно или нет, тут не принципиально для калия). В результате, натрий лавиной обрушивается в клетку, калий тоже быстро выходит из клетки, но на фоне лавины натрия это незаметно, поэтому мембранный потенциал быстро и сильно изменяется вплоть до положительных значений до пика ПД.
Быстрая реполяризация. В быструю деполяризацию мы наблюдали открытие всех воротных натриевых и калиевых каналов, сильный выход калия и ещё более сильный вход натрия, за счёт последнего график шёл вверх. Но, когда был достигнут пик ПД, все натриевые каналы инактивировались (закрылись медленные ворота), ток натрия в клетку резко прекратился, а вот ток калия из клетки вообще-то не прекращался. Поскольку теперь нет притока натрия в клетку, который компенсировал отток калия из клетки, клетка очень быстро теряет «плюсы», поэтому мы наблюдаем резкое изменение мембранного потенциала в отрицательную сторону (график идёт вниз). Причём обратите внимание: пока потенциал положителен, все натриевые каналы именно инактивированы, а не закрыты.

Но по ходу быстрой реполяризации, когда мембранный потенциал принимает отрицательные значения, натриевые каналы из состояния инактивированных (быстрые ворота открыты, медленные закрыты) переходят постепенно в состояние закрытых (быстрые закрты, медленные открыты). Чем более отрицательный мембранный потенциал, тем больше натриевых каналов становятся закрытыми, а концу быстрой реполяризации (при достижении Е-критического) все натриевые каналы становятся закрытыми и готовыми к работе (напоминаю, что инактивированные каналы непригодны к работе, ибо их нельзя открыть).

Следовая деполяризация. Мембранный потенциал возрастает (в отрицательную сторону, т.е. график идёт вниз), однако уже не так быстро – часть калиевых каналов начинает закрываться (на рисунке я закрыл один канал).

Следовая гиперполяризация. Мембранный потенциал уже достиг Е-критического, но часть воротных каналов для калия ещё открыты, поэтому потенциал продолжает расти (график вниз), но последние воротные каналы для калия закрываются постепенно и потенциал восстанавливается до исходного уровня.

Теперь вы точно знаете всё об общем механизме возбуждения/ответа/ПД. А как его регистрируют? МПП мы регистрировали микровольтметром, но с ПД так не получится. Почему? А потому, что МПП (вспомните график) не изменяется во времени, поэтому значение на микровольтметре постоянное, а ПД – это быстрое колебание МПП по определению, т.е. на микровольтметре мы бы максимум заметили бы, что показания прибора колеблются, поэтому ПД замеряют с помощью осциллографа. Осциллограф, как микровольтметр, измеряет мембранный потенциал, но ещё и фиксирует, как тот меняется во времени, показывая время, амплитуду, количество, частоту ПД.
Какое практическое значение имеет регистрация ПД? Мышечная и нервная ткань относятся к возбудимым, они в процессе функционирования постоянно генерируют ПД, поэтому ПД могут характеризовать работу этих тканей. В неврологии и психиатрии, например, очень распространена такая процедура, как электроэнцефалография ЭЭГ, которая регистрирует электрические потенциалы в разных участках мозга и может указывать на нарушения его работы.
Есть два понятия, неразрывно связанных с возбуждением (запомните, пожалуйста, что клеточный ответ = ПД = возбуждение), это лабильность и проводимость. Проводимость – способность клетки распространять волну возбуждения вдоль мембраны. Как это происходит? Мы с вами говорили о натриевых каналах, потенциал зависимых натриевых каналах. Если в одном участке клетки мы вызвали ПД, то в этом участке потенциал на короткое время становится положительным, вокруг возбуждённого участка находятся невозбуждённые участки, чьи натриевые каналы включаются этим самым возбуждением, ведь они реагируют на этот высокий потенциал. Т.е. возбуждение включает натриевые (ну и калиевые) каналы в невозбуждённых участках мембраны, потому что эти каналы потенциал зависимые, а возбуждение – тот самый потенциал, от которого они зависят. Получается, что возбуждённый участок мембраны возбуждает невозбуждённые участки и распространяется так по всей мембране. Схематично изобразил это на рисунке ниже. Плюсы и минусы показывают заряд мембраны (снаружи в покое он положительный), ПД – как известно, быстрое колебание МПП с изменой знака заряда мембраны, поэтому на рисунке ПД можно узнать по изменившемуся заряду мембраны. Смотрите: электростимулятором мы вызвали ПД, который начал возбуждать близлежащие невозбуждённые участки мембраны, а те начали возбуждать другие и так пошла волна возбуждения. Вот это проводимость, определение простое – постарайтесь запомнить, но лучше понять и научиться формулировать определения самостоятельно, исходя из смысла.

Почему волна возбуждения не возвращается обратно? Это станет вам понятно, когда мы будем подробно разбирать тему «возбудимость», а пока лишь скажу, что после пика ПД некоторое время все, а потом лишь часть натриевых каналов инактивированы, т.е. включить их нереально и вызвать новый ПД тоже нереально. А что же такое лабильность? Это время, в которое может развернуться ПД, то есть сколько длится ПД, измеряется в милисекундах, хотя есть и другое определение: лабильность – самое короткое возможное время через между двумя стиумалами, при котором оба стимула вызовут ПД. Грубо говоря, вот вы дали стимул – начал развиваться ПД, даёте новый стимул через 0,01 мс, но нового ПД нет, даёте стимул через 0,05 мс – опять ничего, даёте стимул через 0,1 мс – ничего, даёте стимул через 0,5 мс – ничего, через 1 мс – ничего, через 1,2 мс – есть новый ПД, значит, лабильность составила 1,2 мс. На теме «возбудимость» мы ещё вернёмся к понятию лабильности и там оно станет понятнее. Кстати из лабильности можно вывести понятие максимальной частоты, для этого одну секунду (1000 мс) надо разделить на лабильность в милисекундах. Например, нейрон может возбуждаться каждую милисекунду, лабильность = 1 мс, значит за одну секунду он может возбудиться 1000 раз – это частота. Частота измеряется в герцах, 1 Гц = 1 раз/с. То есть лабильность 1 мс, а частота максимальная 1000 Гц; лабильность 5 мс, тогда максимальная частота, с которой может возбуждаться ткань/клетка, равна 200 Гц (1000 : 5 = 200).
Вспомните, что мы говорили о фазах ПД – медленная деполяризация не входит в состав ПД или считается нулевой фазой, потому что она не всегда приводит к формированию ПД. Чтобы медленная деполяризация привела к ПД, необходимо, чтобы мембранный потенциал от Е-нулевого дошёл до Е-критического – такое расстояние на графике называют дельта-Е или пороговый потенциал. Пороговый потенциал – величина, которую должна преодолеть медленная деполяризация, чтобы вызвать ПД. Если раздражитель способен вызвать деполяризацию, достигающую 100% от порогового потенциала, то появляется ПД, а раздражитель называется пороговым или сверхпороговым (порог раздражителя – минимальная сила, которую должен сообщить раздражитель, чтобы вызвать возбуждение; если раздражитель соответствует порогу, он пороговый, а если он сообщает больше силы, то он сверхпороговый; пороговый потенциал и порог раздражителя – разные вещи), а вот подпороговые раздражители вызывают деполяризацию, которая не достигает 100% от порогового потенциала, угасает и не вызывает ПД. Если деполяризация достигает величины меньше 50% порогового потенциала, то такую деполяризацию называют электротоном, если выше 50%, но ниже 100%, то это локальный ответ. Электротоны призваны регулировать возбудимость клеток. Каким образом? Вспомните, как развивается ПД во время медленной деполяризации: по очереди открываются натриевые каналы, впуская натрий в клетку и уменьшая потенциал (двигали график вверх) до Е-критического. Что делают электротоны? Если на клетку действует подпороговый раздражитель довольно долго, то он открывает только некоторые натриевые каналы, которые спустя какое-то время инактивируются и не могут включиться. Когда на клетку действует пороговый и сверхпороговый раздражители, медленная деполяризация протекает довольно быстро и все открывшиеся натриевые каналы не успевают инактивироваться – всё происходит слишком быстро. А подпороговый раздражитель слабый, ПД не вызывает, медленная деполяризация протекает реально медленно, поэтому некоторые натриевые каналы, которые были включены, успевают инактивироваться. Почему падает возбудимость в таком случае? Ну допустим для вызова ПД нужно открыть 20 натриевых каналов, с этим справится пороговый раздражитель, эффекта от этих каналов хватит, чтобы достичь Е-критического и запустить ПД, но из-за длительного действия подпорогового раздражителя образуется электротон и 4 канала из 20 инактивируются, и когда на клетку подействует пороговый раздражитель, он сможет включить только 16 каналов, а этого не хватит для вызова ПД. То есть в норме/покое клетка бы возбудилась от порогового раздражителя, но под действием длительного электротона это не получается, т.е. клетка стала менее возбудима. Но также электротоны могут и повышать возбудимость клеток, если их действие не длительно. Как? Допустим Е-нулевое равно -70 мВ, Е-критическое равно -50 мВ, тогда раздражитель должен приложить много сил, чтобы изменить мембранный потенциал на 20 мВ, а если мы один раздражитель вызовет электротон (маленькую деполяризацию), то потенциал на время станет равным -65 мВ, тогда раздражителю нужно будет изменить потенциал не на 20, а на 15 мВ, т.е. затратить меньше работы для вызова ПД, т.е. ткань/клетка стала более возбудимой. Стимул от основного раздражителя пришёлся в первую часть электротона, когда натриевые каналы не успели инактивироваться. Вот эту тонкую грань между двумя видами действия электротонов, пожалуйста, уловите. Кстати примерно по такому же принципу (длительное воздействие подпорогового раздражителя, долгий электротон) работает электронаркоз. Для чего нужен локальный ответ? Вообще он возникает в синапсах между нейронами и называется ВПСП (возбуждающий постсинаптический потенциал), но это происходит в возбуждающих синапсах, а в тормозных синапсах возникает ТПСП (тормозной постсинаптический потенциал, обратное явление – истинная гиперполяризация, типа деполяризация, только график не вверх, а вниз), нейрон суммирует все, поступившие на него ВПСП и ТПСП, на основе чего принимает решение, формировать ПД на аксоне или нет. Но это всё вам пока рано знать, я просто сказал это, чтобы вы знали, где используется локальный ответ, его особенность заключается в том, что он способен к суммации (с другими локальными ответами, ну или с ТПСП). И электротон, и локальный ответ, в отличии от ПД, не способны к распространению вдоль мембраны (они имеют слишком низкий потенциал, чтобы включать каналы по соседству), хотя локальные ответы частично так не очень далеко, но распространяются, ибо сигнал от синапса на нейроне как-то должен дойти до аксонального холмика, но это уже тонкости, официально мы говорим, что ЭТ и ЛО не распространяются за пределы действия раздражителя, который их вызывает.

Возбудимость.
А теперь наконец поговорим о таком важном понятии, как возбудимость. Напоминаю, что возбудимость – это способность клеток/тканей создавать быстрое колебание МПП с изменой знака заряда мембраны, т.е. генерировать ПД, в ответ на действие раздражителя. Возбудимость определяется параметрами раздражителя и морфофункциональным состоянием мембраны клеток. Разберёмся в этом подробнее и начнём с параметров раздражителя, их три: сила, время действия, градиент нарастания силы во времени.
Сила и время действия рассматриваются исключительно совместно, так как между ними есть обратная зависимость, что можно наблюдать на графике. Вверх отмечаем силу действия раздражителя в милиамперах, а вправо – время в милисекундах.

Риабаза – минимальная сила, способная вызвать возбуждение. Иначе риабазу называют абсолютным порогом силы (если пересечь этот порог силы, то сколько времени не раздражай мембрану, а возбуждение не получишь). Время, которое требуется риабазе, чтобы вызвать возбуждение, называют полезным временем. Время, которое потребуется, чтобы вызвать возбуждение при действии силы в две риабазы, называется хроноксией. Но ещё существует и так называемый абсолютный порог времени, т.е. если раздражитель действует меньше этого времени, какой бы силой он не обладал, возбуждение он не вызовет. Любая точка на кривой, которую я начертил, означает возбуждение, соответствующее определённой силе и времени, продлять эту линию за точки, которыми я её ограничил, категорически запрещено, ибо две эти точки обозначают пределы, за которыми возбуждение невозможно. Поэтому график чертите именно так. Что нам даёт этот график? Во-первых, он говорит о том, что для вызова возбуждения нужна определённая достаточная сила (порог силы), во-вторых, он говорит о том, что существует такое время действия раздражителя, в течение которого ПД точно не возникнет (порог времени), поэтому если шприц воткнуть в ягодицу достаточно быстро, то пациент практически не чувствует укола, в-третьих, между силой и временем существует обратная зависимость (чем выше сила раздражителя, тем меньше ему требуется времени, чтобы вызвать возбуждение), в-четвёртых, хроноксия – мера возбудимости. Подробнее про хроноксию. Почему мы используем не полезное время, а хроноксию? Полезное время – предельная точка, показывающая границы графика, а вот хроноксия – точка «внутри» графика и является более информативной, от неё зависит степень изгиба графика. Если вы не физмат, то могу дать определение иначе. У двух клеток одинаковые пороги силы и времени, но хроноксия разная. У первой ткани хроноксия 15 мс, а у второй 20 мс, т.е. при действии на обе клетки одной и той же силой (две риабазы) первая клетка возбуждается быстрее второй, поэтому возбудимость первой клетки выше. Хроноксия обратно пропорциональна возбудимости.
Третий параметр раздражителя – градиент нарастания силы во времени. Как это понимать? Ну вот клетка находится в покое, сила, действующая на неё, равна 0 мА, потому, что никакой раздражитель на неё не действует, но потом появляется какой-то раздражитель с силой 10 мА, но сразу ли на клетку обрушиваются все эти 10 милиампер или они делают это в течение какого-то времени постепенно? Часто бывает второе, т.е. сила достигает своего максимума только через какое-то время, т.е. она постепенно нарастает в течение времени, следовательно, существует скорость нарастания силы в течение времени, мы называем эту скорость градиентом нарастания силы во времени. Чем выше градиент, тем выше возбудимость. Почему? Ну допустим, градиент очень высокий, тогда в медленную деполяризацию открылись натриевые каналы и не успели инактивироваться, как началась быстрая деполяризация. А если градиент не высокий, то натриевые каналы в медленную деполяризацию открываются так медленно, что пока откроются одни, другие уже успеют инактивироваться, тогда приток натрия в клетку ограничивается, а возбудимость падает (это как при длительном электротоне). Возбудимость растёт за счёт того, что Е-критическое изменяется (из-за инактивации натриевых каналов) и при низком градиенте нарастания силы во времени может быть никогда недостигнуто! Пример: если поставить иглой проткнуть кожу ягодиц, а затем очень медленно давить на иглу, наращивая силу нажатия очень постепенно, пациент не почувствует, что вы вгоняете иглу в глубь. Кстати есть такая фишка: чем выше сила, тем быстрее проходит медленная деполяризация. Как продемонстрировать это на графике? См. ниже.

Сила раздражителя здесь одинаковая, а вот скорость её нарастания разная.
Теперь поговорим о параметрах мембраны. Тут всего один параметр – морфофункциональная целостность. Что это значит? Морфологическая целостность означает, что в мембране нет дыр, все белки на месте и тд – все структуры мембраны в норме. Функциональная целостность означает, что все структуры готовы к работе, т.е. каналы для натрия не забиты анестетиками, в клетке имеется АТФ для работы натрий-калиевой-АТФазы, рецепторы каналов не забиты никакими ядами и тд.
Если нарушить любой из параметров (переступить пороги времени, силы, сделать слишком низкий градиент нарастания силы во времени, нарушить структурно-функциональную целостность мембраны), то возбуждения не будет.
Итак, мы вспомнили, что такое возбудимость, обсудили условия, от которых она зависит (параметры возбудимости или условия возникновения возбуждения), а теперь обсудим меры возбудимости: порог раздражителя, хроноксия, пороговый потенциал, лабильность, частота. Порог раздражителя и хроноксия относятся к параметрам раздражителя, а пороговый потенциал, лабильность и частота (максимальная частота возбуждения) относятся к параметрам возбудимой системы (зависят от свойств мембраны).
Порог раздражителя – минимальная сила, которую должен сообщить раздражитель возбудимой системе, чтобы в той возникло возбуждение (измеряется в милиамперах). Хроноксия – время действия раздражителя, которое потребуется на формирование ПД при действии на систему двух риабаз (измеряется в милисекундах). Пороговый потенциал – величина, которую должна преодолеть деполяризация, чтобы достигнуть Е-критическое (измеряется в миливольтах). Лабильность – время, в течение которого разворачивается единичный ПД (измеряется в милисекундах). Максимальная частота возбуждения – максимально возможное количество ПД, генерируемых клеткой за одну секунду (измеряется в герцах), величина, производная от лабильности. Что касается частоты действия раздражителя, то она не всегда соответствует частоте возбуждений клетки по ряду причин (у клетки есть предел частоте, а раздражитель может быть выше этого предела; чем выше частота раздражителя, тем короче время его действия, при слишком высокой частоте мы имеем слишком короткое время действия раздражителя, которое может пересечь абсолютный предел времени и не сможет вызывать ПД).
А теперь рассмотрим очень важный и интересный вопрос, который прояснит некоторые темы, разобранные выше, в частности вопросы лабильности и частоты. Мы разберём график изменения возбудимости в процессе возбуждения.

Примем, что возбудимость клетки в покое равна 100%. Во время медленной деполяризации возбудимость растёт. Почему? Потому что мембранный потенциал по мере роста медленной деполяризации всё ближе и ближе к Е-критическому, т.е. пороговый потенциал уменьшается, следовательно, возбудимость растёт. Фаза медленной деполяризации соответствует фазе первичной экзальтации (повышенная возбудимость, возбуждение могут вызвать даже подпороговые раздражители). Затем начинается быстрая деполяризация – возбудимость падает до нуля мгновенно. Почему? Потому, что в быструю деполяризацию открываются все натриевые каналы, т.е. возбуждать больше нечего – всё уже возбуждено. Когда наступает фаза быстрой реполяризации, все натриевые каналы оказываются инактивированными, включить их невозможно – опять некого возбуждать, возбудимость на нуле, но так будет до тех пор, пока потенциал не достигнет нуля, после этого натриевые каналы начнут переходить в состояние закрытых, а закрытые каналы можно открыть, т.е. будет, что возбуждать – возбудимость станет расти. Выходит, что в фазу быстрой деполяризации и в фазу быстрой реполяризации (конкретно та её часть, где потенциал больше нуля) возбудимость равна нулю – фаза абсолютной рефрактерности (ничто не может возбудить клетку). Далее в оставшуюся часть фазы быстрой реполяризации (потенциал ниже нуля) натриевые каналы переходят в закрытое состояние – есть, кого возбуждать, возбудимость растёт. Эта часть фазы быстрой реполяризации соответствует фаза относительной рефрактерности (возбуждение возможно, но только при действии сверхпороговых раздражителей, так как не все натриевые каналы в деле). Фаза следовой деполяризации соответствует фазе вторичной экзальтации (повышенная возбудимость, возбуждение способны вызвать подпороговые раздражители), механизм здесь тот же, что и при первичной экзальтации, только форма графика иная. Почему? В точке Е-критическое возбудимость должна достигать пика, но этого не происходит в связи с обильным выходом калия из клетки (в начале следовой деполяризации калий выходит быстрее, чем в начале быстрой деполяризации, поэтому в первый раз в точке Е-критическое у нас очень высокая возбудимость, а во второй – не очень). Фаза следовой гиперполяризации соответствует фазе субнормальной возбудимости (возбудимость снова невозможна), так как мембранный потенциал растёт в отрицательную сторону, увеличивая тем самым пороговый потенциал (типа был 20 мВ, а возрастает аж до 40 мВ – видно на графике).
Теперь мы разобрались в том, как изменяется возбудимость в процессе возбуждения, но отсюда можно вынести пару моментов, которые важно знать. Во-первых, лабильность. Лабильность равна фазе абсолютной рефрактерности. Если после очередного стимула (который пришёлся в точку окончания абсолютной рефрактерности) пустить новый стимул, выждав время меньшее, чем лабильность, то вы попадёте в абсолютную рефрактерность и нового ПД не возникнет, а если вы выждите время, равное лабильности, то снова получите ПД. Лабильность – самое минимально возможное время разворачивания единичного ПД, поэтому лабильность определяет максимально возможную частоту возбуждения клетки – быстрее невозможно. Во-вторых, частота самого раздражителя. Подбирая разную частоту раздражителя, вы можете заметить, что ткань/клетка ведёт себя совершенно по-разному. Чем выше частота раздражителя, тем ближе по времени друг к другу стимулы/импульсы. Небольшая частота: два импульса дают два ПД с равной амплитудой (первый импульс вызывает ПД, второй импульс действует на клетку, когда та уже успела прийти в состояние покоя – нормальная возбудимость, и вызывает полноценный ПД). Частота побольше: два импульса дают один ПД (первый импульс вызывает полноценный ПД, а второй попадает в фазу субнормальной возбудимости, поэтому не может вызвать ПД). Ещё более высокая частота: два импульса дают два ПД с равной амплитудой (первый импульс вызывает полноценный ПД, второй импульс попадает в фазу вторичной экзальтации и легко вызывает полноценный ПД). Очень высокая частота: два импульса дают два ПД, но у второго амплитуда меньше (первый импульс вызывает полноценный ПД, а второй импульс попадает в фазу относительной рефрактерности (будем думать, что раздражитель сверхпороговый) и вызывает ПД, но с меньшей амплитудой, если раздражитель будет и дальше работать с такой частотой, то амплитуды будут становиться всё меньше, пока ПД просто не пропадут). Слишком высокая частота: два импульса дают один ПД (первый импульс вызывает полноценный ПД, а второй импульс приходится в фазу абсолютной рефрактерности и не способен вызвать ПД).
Ответная реакция при возбуждении.
Теперь поговорим о факторах, которые определяют характер ответной реакции биосистемы на раздражение. Эти факторы представлены параметрами раздражителя и морфофункциональным состоянием мембраны. Понятно, что при нарушении морфофункционального состояния степень ответа биосистемы снижается или и вовсе отсутствует. Теперь о параметрах раздражителя. Если действовать с низким градиентом, то ответ ослабляется или и вовсе отсутствует (вспомните графики ПД для разных градиентов – результат инактивации натриевых каналов). Если действовать недостаточное количество времени, то ответа просто не будет (медленной деполяризации требуется время, чтобы дойти до Е-критического). Но самым важным для нас является фактор/параметр силы раздражителя. Как зависит характер ответной реакции биосистемы от силы раздражителя? У тканей и клеток это две разные истории – разберём каждую.
Для клетки.
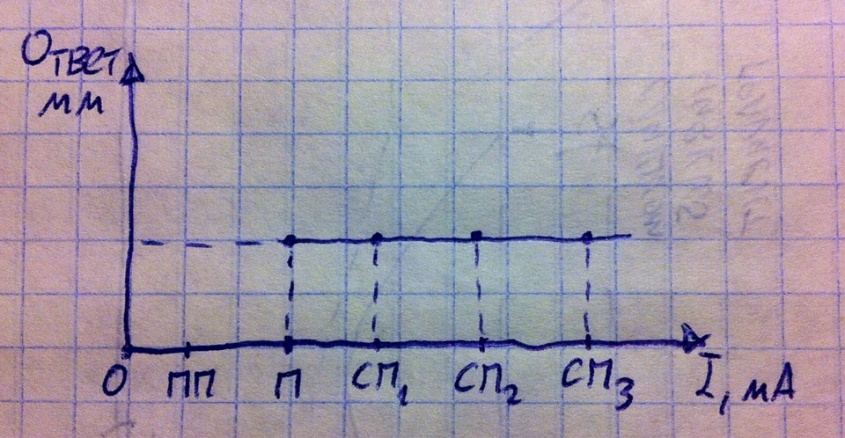
Исследуемой клеткой пускай будет мышечное волокно – его ответной реакцией на раздражение (точнее на возбуждение) является сокращение, т.е. уменьшение длины, которое на практике можно измерить в милиметрах; сила раздражителя измеряется в милиамперах. Для клетки действует закон «всё или ничего». То есть ответа либо нет, либо он максимален. То есть подпороговый раздражитель не вызывает ответной реакции (на то он и подпороговый), пороговый вызывает максимальный ответ, сверхпороговые тоже вызывают максимальный ответ. Почему так? А всё потому, что во время ПД в клетке открываются все натриевые каналы уже при действии порогового раздражителя, от того, что вы будите повышать силу раздражителя толку нет – все каналы уже открыты, открывать больше нечего. Обратите внимание: на графике есть одна предельная точка, которая соответствует пороговому раздражителю, до неё возбуждение и ответ невозможны (то есть продлять график за эту точку нельзя – ошибка), а вот после этой точки возбуждение и ответ будут всегда – продлевайте в ту сторону график, сколько хотите.
Для тканей.

Для тканей действует закон «силовых отношений». Как это? Подпороговые раздражители не вызывают ответа (понятно). Пороговый раздражитель вызывает минимальный ответ. Максимальный раздражитель вызывает максимальный ответ. Субмаксимальные раздражители (больше порогового, но меньше максимального) вызывают ответы разной силы промежуточных значений. Сверхмаксимальные раздражители вызывают такой же ответ, как и максимальный раздражитель. Откуда такая странная закономерность? Дело в том, что ткань (пускай это будет скелетная мускулатура) состоит из множества клеток (мышечных волокон; вообще-то это не клетки, а симпласт, но это не важно), и эти клетки обладают разной возбудимостью! Т.е. у разных клеток разные пороги. И что получается? Вот подействовали пороговым раздражителем – возбудилось 10% клеток – мышца сократилась на 10 мм. Подействовали субмаксимальным раздражителем – возбудили 40% клеток – мышца сократилась на 40 мм. Действуете максимальным раздражителем – возбуждается 100% клеток – мышца сокращается на 100 мм. Действуете сверхмаксимальным раздражителем – всё равно возбудится 100% и сокращение на 100 мм, потому что больше просто невозможно. То есть тут ответ может зависеть от силы (в пределах между пороговой и максимальной точками) в связи с тем, что в ткани клетки имеют разный уровень возбудимости и чем выше сила, тем больше клеток возбудится и ответит и так до тех пор, пока не возбудятся все клетки.
Физиология нервных проводников и мышечных волокон.
А теперь приступим к частной физиологии нервных проводников и мышц.
Начнём с мышц (скелетная мускулатура). Что из себя представляет отдельно взятая мышца? Это совокупность однонаправленных мышечных волокон (так называется мышечная клетка, точнее симпласт, а не клетка), собранных в пучки и обмотанных соединительной тканью, плюс у мышцы есть соединительнотканные сухожилия и определённая иннервация, но главная часть мышц – мышечные волокна, именно они составляют большую часть всей мышцы и реализуют главное свойство мышц – сократимость. Давайте скажем пару слов о мышечных волокнах. Они бывают разные: есть интрафузальные (служат больше не для сокращения мышц, а вспомогательной клеткой для рецептора растяжения мышц) и экстрафузальные (вот их главная задача – сокращаться), экстрафузальные подразделяют на тонические (красные, сокращаются медленно, обладают низкой лабильностью, зато они способны сокращаться очень долго, для этого у них есть большие запасы гликогена и миоглобина, который и даёт красный цвет) и фазические (белые, сокращаются быстро, имеют высокую лабильность (высокую в смысле короткую, типа очень быстро ПД развивается), они имеют меньше запасающих веществ и работают недолго). Однако немного некорректно говорить, что мышца состоит из мышечных волокон – правильнее будет сказать, что мышца состоит из двигательных единиц. Двигательная единица – совокупность мышечных волокон, иннервируемых одним мотонейроном (все мышечные волокна в составе двигательной единицы сокращаются одновременно или не сокращаются одновременно, потому что иннервируются общим нейроном). Исходя из классификации мышечных волокон, строится классификация мышц (один из вариантов их классификации): сильные и быстрые мышцы. В сильных мышцах преобладают медленные/красные мышечные волокна, а в быстрых мышцах преобладают быстрые/белые мышечные волокна.
Для чего нам вообще нужны скелетные мышцы? Во-первых, для движений. Движения бывают позными (длительное сокращение некоторых мышц позволяет нам долгое время поддерживать определённую ориентацию тела в пространстве – позу) и целенаправленными (классические движение, типа ходить ногами, махать руками и тд). Во-вторых, мышцы необходимы для термогенеза. Как это понимать? Мышечное сокращение имеет низкий коэффициент полезного действия, т.е. мышцы тратят на сокращение много энергии, но большая часть этой энергии рассеивается в форме тепла, которое согревает организм. Именно поэтому после длительной пробежки вам становится жарко, другой пример мышечного термогенеза: когда вы стоите на холодной улице, вы начинаете дрожать – это быстрое сокращение ваших мышц, призванное хоть как-то вас согреть. В-третьих, мышцы являются аварийным источником аминокислот в организме, так как они переполнены белками (если печень решает устроить глюконеогенез, то ей нужны аминокислоты, которые она забирает из крови и из белков крови и мышц, есть и другие случаи, когда белки мышц приходится гидролизовать, но это уже вопросы биохимии). Какие свойства присущи мышечной ткани? Это возбудимость, проводимость и сократимость. Возбудимость и сократимость мы уже подробно изучили в темах выше, а вот сократимость – новая тема, давайте разберёмся в ней подробнее.
Сократимость – способность клетки уменьшать свою длину (сжиматься), требующая затрат энергии и активируемая потенциалом действия. Прежде, чем начать обсуждать механизм сокращения, надо немного вспомнить морфологию мышечных волокон (не всю, а только моменты, важны с точки зрения физиологии). Большую часть мышечного волокна занимают миофибриллы – сложные белковые комплексы, способные к сокращению, эти комплексы во множестве мест прикрепляются к мембране клетки, чтобы при сокращении миофибрилл сама клетка тоже сокращалась, двигаясь вслед за миофибриллами. Также в мышечных волокнах есть Т-системы – впячивания мембран, около которых располагается саркоплазматический ретикулум СПР, в котором содержится большое количество ионов кальция, СПР имеет каналы с райодиновыми рецепторами, которые по команде открываются и выпускают кальций в саркоплазму (цитоплазму), и кальциевая АТФаза, которая закачивает кальций обратно. Теперь перейдём к самим миофибриллам. Миофибриллы состоят из чередования миозиновых и актиновых «компонентов», которые я обозначил на рисунке красным и синим соответственно. Актиновый компонент. Имеется Z-линия, которая крепится к саркоплазматической (цитоплазматической) мембране для фиксации миофибрил, к этой полоске перпендикулярно присоединяются сложные комплексы из множества белков актина, тропонина и тропомиозина. Миозиновый компонент. Имеется аналогичная М-линия (только она не прикрепляется на мембране), на ней перпендикулярно расположены сложные комплексы из миозина. Миозиновый компонент для стабильности присоединяется к Z-линии с помощью титина и сердина. Актиновый и миозиновый компоненты проникают друг в друга и способны скользить друг относительно друга, т.е. могут проникать друг в друга ещё глубже, укорачивая тем самым миофибриллу. Участок миофибриллы, заключённый между двумя соседними Z-линиями, называется сакромером, он является структурно-функциональной единицей миофибрилл.

Вот мы и обсудили всю важную морфологию. Если что, на мембране отметил заряды и стимул, чтобы изобразить ПД и его распространение. Теперь к биохимическим аспектам, без которых механизм сокращения будет понять сложно. Главные белки в миофибриллах: миозин, актин, тропомиозин, тропонин.
Миозин имеет 6 протомеров: 2 головки, 2 шарнирных участка, 2 тяжёлые цепи. Актин – мономерный белок, имеющий 2 кластера для связывания с другими актинами (это чтобы множество отдельных актинов объединились друг с другом в длинную цепь белков, мономерный белок становится полимерным) и один центр связывания с миозином для образования актин-миозиновых мостиков. У актина есть G-форма (одиночный актин) и F-форма (полимеризованный актин). Тропомиозин – белок, состоящий из двух протомеров, этот белок полимеризуется, образуя двойную спираль. Тропомиозин следует вдоль актина и закрывает его центры связывания с миозином. Тропонин состоит из трёх протомеров (TnI, TnC, TnT). Тропонин располагается между актином и тропомиозином, при активации тропонина, тот сдвигает тропомиозин, который открывает центры связывания актина с миозином, делая сокращение возможным. Т.е. сокращением занимаются актин и миозин, а тропомиозин и тропонин больше выполняют регуляторную функцию.

Ну а теперь наконец-то можно приступить к механизму мышечного сокращения. Оно активируется ПД, возникающем в нервно-мышечном синапсе, ПД распространяется по мембране и активирует аденилатциклазную систему (подробности её работы вы можете узнать в курсе биохимии в теме гормональной регуляции, но сейчас это не принципиально), которая приводит к образованию цАМФ, также ПД перезаряжает мембраны в Т-системах, это улавливают райодиновые рецепторы саркоплазматического ретикулума и открывают каналы для кальция, тот выходит в саркоплазму (если что, концентрация кальция в СПР в тысячи раз выше, чем в саркоплазме в покое). В итоге, у нас образовались цАМФ и кальций, они поступают к тропонину и связываются с его субъединицами: сначала цАМФ соединяется с TnI, конформация белка изменится, после этого кальций соединится с TnC, тогда конформация тропонина изменится ещё больше и TnT заставит тропомиозин подвинуться и открыть центры связывания актина с миозином. Теперь сокращение может начаться, а до этого всё был «подготовительный» этап (неофициальное название, официально его называют латентным периодом, но к этому мы ещё вернёмся). Миозин на своей головке имеет частично гидролизованный АТФ (комплекс АДФ и ортофосфата, который пока не отдал свою энергию), миозин самопроизвольно подтягивается к актину и встаёт своей головкой в его центр связывания – образуется актин-миозиновый мостик. После образования этого мостика АТФ гидролизуется полностью, выделяя энергию, часть которой идёт на конформационные изменения шарнирного участка миозина – участок этот начинает сгибаться в месте крепления с тяжёлой цепью (был между ними угол 90 градусов, а стал 45), в результате таких подтягивающих движений миозин движется относительно актина. В актин-миозиновом мостике всё ещё остаётся АДФ после полного гидролиза АТФ, а для продолжения сокращения нужно разрушить мостик, чтобы потом могли образоваться новые мостики. На место АДФ встаёт АТФ и частично гидролизуется (без отдачи энергии превращается в комплекс АДФ и ортофосфата), мостик разрушается, шарнирный участок миозина восстанавливает конформацию (угол с 45 градусов возрастает до 90), оказываясь вместе с головкой напротив уже нового актина. Дальше всё повторяется. Так происходит сокращение.

А как происходит расслабление? Смотрите: ПД нужен был, чтобы в саркоплазме появились кальций и цАМФ, которые активируют миофибриллу через тропонин, значит, чтобы вызвать расслабление, нужно избавиться от кальция и цАМФ. Как это происходит? ПД прекратился, эти вещества больше не поступают в саркоплазму, теперь они только уходят из неё: цАМФ с помощью дифосфоэстеразы превращается в просто АМФ, а кальций закачивается обратно в саркоплазматический ретикулум кальциевой АТФазой. Кальций и цАМФ больше не связываются с тропонином, он восстанавливает свою конформацию, возвращая тропомиозин на место, тот закрывает центры связывания актина и тот более не может образовывать мостики с миозином – сокращение более невозможно, происходит расслабление. Прошу обратить внимание, что и сокращение, и расслабление тратят много АТФ прямым или косвенным способом. В латентном периоде мы образуем цАМФ, но делаем-то мы их из АТФ. В период сокращения АТФ тратится непосредственно для движения миозина относительно актина – АТФ отдаёт свою энергию на конформацию шарнирного участка. В период расслабления кальциевая АТФаза тратит АТФ для первично активного транспорта кальция из саркоплазмы в СПР, плюс АТФ по факту нужен для разрушения актион-миозиновых мостиков, но это можно отнести и к сокращению, и к расслаблению. На этом всё о механизме мышечного сокращения. Для любознательных: после смерти АТФ перестаёт вырабатываться, поэтому расслабление поддерживать не получается, а тем временем мембраны не работают – в клетку проникает кальций, вызывая активацию актина, получается, что актин-миозиновые мостики не разрушаются – мышцы сохраняют ту длину, которую им придали изначально – трупное окоченение.
Я тут говорил о некой латентной фазе, сокращении, расслаблении. Скелетные мышцы работают в три фазы: латентную, сокращение и расслабление. Все они отмечаются на графике одиночного мышечного сокращения.

С момента подачи ПД и до наступления сокращения период именуется латентным. Почему? Потому что сокращение уже запущено, но требуется время, чтобы его реализовать (пока возбуждение распространится по мембране, пока выйдет достаточное количество кальция из СПР, пока аденилатциклазная система успеет наделать цАМФ, пока тропонин сместит тропомиозин – всё это время). Сам ПД занимает не весь латентный период, а небольшую его часть, т.е. мышечное волокно большую часть латентного периода, весь период сокращения и весь период расслабления обладает нормальной возбудимостью, а это значит, что можно вызвать новое сокращение даже тогда, когда старое сокращение ещё не прекратилось! Так обычно и происходит – в нашем организме практически не бывает одиночных сокращений, только тетанические (много подряд). Тетанус (тетанические сокращения) бывает гладким и зубчатым. Если на мышечное волокно действует стимул, когда то находится в фазе расслабления, то мы наблюдаем зубчатый тетанус (график похож на зубцы и канавки). А если стимулы подаётся на мышечное волокно, когда то находится в фазе сокращения, наблюдаем гладкий тетанус (графики сокращений расположены так близко, что поверхность общего графика почти гладкая, во всяком случае по сравнению с зубчатым тетанусом).

Итак, виды мышечного сокращения: одиночные и тетанические, а тетанические подразделяются на гладкий и зубчатый тетанус.
А какие есть типы мышечных сокращений? Изометрическое и изотоническое. Изо – постоянность, метр – длина (в данном случае), т.е. при изометрическом сокращении длина мышцы/волокон остаётся постоянной, а вот напряжение изменяется. Как это? Ну вот стоит грузовик, вы пытаетесь его поднять, вы прикладываете усилие в 50% от своего максимума - не получается, вы повышаете силу до 90% - грузовик всё там же, не сдвинулся с места, соответственно и ваши руки остались там, где и были, длина мышц, которые пытаются согнуть ваши руки, чтобы те подняли грузовик, не изменяется (изометрия), а вот напряжение вы изменяли (с 50% на 90%). Изотоническое напряжение предполагает постоянный уровень напряжения мышц/волокон, а вот длина их изменяется. Как это? Вы поднимаете гирю, постоянно прилагаю одно и то же усилие – напряжение постоянно, а вот длина мышцы уменьшается, и она сгибает руку в локтевом суставе. Пожалуйста, ни в коем случае не путайте виды (одиночное и тетаническое) и типы (изометрическое и изотоническое) мышечных сокращений!
Теперь в теме частной физиологии мышц осталось разобрать несколько второстепенных, но всё-таки важных вопросов: от чего зависит сила мышц, утомление, особенности гладких мышц.
Начнём с силы мышц. Ответственными за сокращение в мышцах/волокнах являются миофибриллы. Во время длительных тренировок мышечные волокна гипертрофируются, т.е. растут в объёме за счёт увеличения числа миофибрилл, но также увеличивается число митохондрий и миоглобина (запас кислорода). Таким образом, первый фактор, определяющий силу мышц – это состав мышечных волокон (количество миофибрилл и миоглобина). Но есть и второй фактор – нервный. Не забывайте, что все сокращения скелетной мускулатуры вызываются сигналами от нервной системы. Если у вас есть какие-то проблемы с её деятельностью (патологические или нормальные физиологические состояния), то она будет посылать менее интенсивные импульсы на мышцы и те будут слабее сокращаться.
Утомление. Если говорить по-простому, то утомление – это прекращение работы системы в связи с определёнными затруднениями в её функционировании (как правило, что-то истощается). Как я уже сказал, деятельность мышц зависит от самих мышц и от нервной системы, активирующей их. Утомляться могут нервные центры, нерв, нервно-мышечный синапс, мышечное волокно. Нервные центры представлены динамической совокупностью синаптически связанных нейронов, регулирующих определённые физиологические процессы и функции, но главное, что это нейроны с синапсами, в синапсах есть небольшой запас нейромедиаторов, если нейроны слишком часто используют синапс, то запас нейромедиаторов истощается и синапс больше не работает, а синапс – средство связи нейронов, без него нейроны не сообщаются друг с другом и работа нервного центра угасает. Нейромедиаторы потом, конечно, синтезируются новые, но это процесс не быстрый. В общем нервные центры при чрезмерно интенсивной работе утомляются из-за истощения запасов нейромедиаторов. По той же причине утомляется нервно-мышечный синапс, вот только у него это утомление наступает гораздо быстрее. Утомить нерв очень сложно, в нерве находятся аксоны, проводящие ПД от тела к синапсу – им нужен только АТФ для работы натрий-калиевой-АТФазы, т.е. нерв не особо требователен, а потому утомляется меньше всех. Утомление мышечных волокон связано с истощением в них запасов гликогена и отчасти из-за истощения кислорода, связанного с миоглобином. Обычно принято говорить об утомлении нерва, нервно-мышечного синапса и мышечного волокна (нервные центры – отдельный вопрос). Самым утомляемым является синапс (медиаторы быстро кончаются, синтезируются небыстро), меньше утомляются мышечные волокна (у них всё-таки приличный запас гликогена и миоглобина), почти неутомимый нерв (ему нужен только АТФ, которого обычно у нейронов предостаточно).
Гладко-мышечные клетки. Они имеют форму веретена. Они не формируют орган-мышцу – они обычно формируют мышечные слои органов и сосудов. Эти клетки обладают автоматией (способность самостоятельно формировать ПД без внешних стимулов), а раз есть автоматия, то они сами вызывают своё сокращение. ГМК (гладко-мышечные клетки) обладают ну очень низкой лабильностью (десятки секунд, а то и минута).
Теперь можно перейти к частной физиологии нервных проводников. Кто относится к этим проводникам? Нервные волокна. Нервное волокно представляет собою отросток нейрона, погружённый в глиальную клетку (отросток - это как металлическая часть провода, а глиальные клетки – изоляция). Отростки нейронов: аксоны (любые) и дендриты (только те, что у спинальных ганглиев, т.е. очень длинные). Нервные волокна бывают миелиновыми и немиелиновыми. Миелиновые волокна отличаются тем, что вокруг нервного отростка намотана мембрана в несколько туров – это позволяет повысить скорость проведения нервного импульса (ПД), однако эта обмотка идёт не вдоль всего волокна – есть регулярные небольшие участки, лишённые миелина – перехваты Ранвье, если бы их не было, то импульсы не могли бы проводиться, плюс нервный отросток лишился бы трофики. У нервных волокон два главных свойства: возбудимость и проводимость. Их главная задача – проведение нервного импульса. Разберёмся чуть подробнее в механизме проведения нервных импульсов.
Для немиелиновых волокон. Вдоль всего нервного отростка расположены натриевые каналы, возбуждение равномерно движется вдоль мембраны – возбуждённые участки мембраны возбуждают прилежащие участки невозбуждённой мембраны, ибо их натриевые каналы реагируют на потенциал действия. На рисунке снизу я схематично показал, как распространяется возбуждение по немиелиновому волокну. Такие волокна медленно проводят сигнал и очень быстро выключаются анестетиками. На рисунке на нервное волокно действуют электростимулятором, вызывая возбуждение. Немиелиновое волокно слева.

У миелиновых волокон в области самого миелина никаких каналов нет, ПД существует только в перехватах Ранвье, но в этих перехватах так много натриевых каналов и импульс такой сильный, что он просто перепрыгивает на соседний перехват – это называется сальтаторное проведение импульса, оно позволяет использовать меньше каналов и проводить сигнал с куда большей скоростью, потому что сигнал перескакивает миелиновый участок быстрее, чем если бы он шёл каналам. Мало того, что миелиновые волокна быстрее проводят сигнал, так они ещё и более устойчивы перед анастезией, ибо сальтаторное проведение порою позволяет перескочить не на соседний перехват Ранвье, а даже через него. На рисунке выше миелиновое волокно справа.
Теперь поговорим о классификации нервных волокон. Их классифицируют в зависимости от наличия миелиновой оболочки и толщины волокна.
Тип волокна |
Скорость проведения (м/с) |
функции |
А-альфа |
70-120 |
Афферентные волокна от мышц |
А-бета |
30-70 |
Афферентные волокна от рецепторов осязания и давления |
А-гамма |
15-30 |
Эфферентные волокна к мышцам |
А-дельта |
12-30 |
Афферентные волокна от болевых и терморецепторов |
В |
3-15 |
Симпатические приганглионарные волокна |
С |
0,5-2 |
Симпатические постганглионарные волокна, афферентация боли |
Если что, афферентные волокна – дендриты (сигнал от рецептора в ЦНС), эфферентные волокна – аксоны (сигнал от тела нейрона к концу аксона на эффектор).
Последний вопрос в теме частной физиологии нервных проводников – законы проведения возбуждения по нерву, их 4.
Закон двустороннего проведения (справедлив для периферических нервов, но не для ЦНС). Если вы возбуждаете нерв не на конце, а где-то на его протяжении, то возбуждение распространяется в обе стороны, ибо возбуждённый участок мембраны возбуждает все прилежащие участки невозбуждённой мембраны, а эти участки имеются по обе стороны от раздражителя, поэтому возбуждение распространяется в обе стороны (логично, кстати на рисунке, демонстрирующем проведение импульса миелиновыми и немиелиновыми волокнами, как раз видно, что возбуждение проводится одинаково в обе стороны). Но почему сигнал не возвращается обратно? Ну потому, что участок мембраны возбуждается, впадает в фазу абсолютной рефрактерности, потом в фазу относительной рефрактерности, т.е. возбудить этот участок снова, чтобы возбуждение пошло в обратную сторону, не получится, т.е. спасибо инактивации натриевых каналов.
Закон анатомической и функциональной целостности. Думаю, здесь всё понятно: если анатомическая целостность нарушена (перерезан нерв), то его работа невозможна, а под функциональной целостностью имеется ввиду, что ничто не мешает работе нерва, например, анестетик мог бы проникнуть в натриевый канал и не дал бы ему работать – возбуждение и проведение невозможно без рабочих натриевых каналов.
Закон изолированного проведения. Сигнал с одного волокна на другое не перескакивает, потому что сигнал по сути идёт по нервному отростку, который завёрнут в глиальную клетку, да ещё и в соединительную ткань может быть дополнительно упакован – отличная изоляция. Если бы изоляции не было, сигнал беспорядочно перескакивал с одних проводников на другие, полностью нарушая адекватную работу нервной системы.
Закон бездекретного проведения. Имеется ввиду, что по ходу волокна амплитуда импульса остаётся неизменной, каким бы длинным волокно ни было. Это связано с тем, что натриевые каналы (которые открываются на Е-критическом) работают по принципу «всё или ничего», т.е. при возбуждении в нервном волокне открываются все натриевые каналы и амплитуда ПД всегда максимальна и, соответственно, постоянна.