

Физиология 4 колок
.pdfСкелетная мышца как активная часть опорнодвигательного аппарата
1. Моторная единица (МЕ) является функциональной единицей скелетной мышцы. МЕ включает в себя группу мышечных волокон и иннервирующий их
мотонейрон. Число мышечных волокон, входящих в состав одной МЕ, варьирует в разных мышцах.
Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на медленные (S-МЕ) и быстрые(F-МЕ). А F-МЕ в свою очередь делят по устойчивости к утомлению на устойчивые к утомлению (FF-МЕ) и быстроутомляемые(FR-МЕ).
Соответствующим образом подразделяют иннервирующие данные МЕ мотонейроны. Существуют S-мотонейроны(S-МН), FF-мотонейроны (F-МН) и FR - мотонейроны(FRМН) S- МЕ характеризуются высоким содержанием белка миоглобина, который способен связывать кислород (О2). FF-МЕ составляют
мышцы, способные выполнять быстрые сокращения без заметного утомления. Волокна FF-ME содержат большое количество митохондрий и способны образовывать АТФ путем окислительного фосфорилирования.
2. https://helpiks.org/7-9968.html
Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется выброс (экзоцитоз) медиатора (лат. mediator — посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках (везикулах), диаметр которых составляет около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание, где заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 тыс. до 10 тыс. молекул химического вещества
Постсинаптическая мембрана (концевая пластинка в нервно-мышечном синапсе) —
это часть клеточной мембраны иннервируемой мышечной клетки, содержащая рецепторы, способные связывать молекулы ацетилхолина. Особенность этой мембраны: множества мелких складок, увеличивающих ее площадь и количество рецепторов на ней до 10—20 млн в одном синапсе.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, в совокупности образующих базальную мембрану, соединяющую пре- и постсинаптическую мембраны.

3. http://vmede.org/sait/?id=Fiziologiya_atlas_kamakin_2010&menu=Fiziologiya_atl as_kamakin_2010&page=6
Скелетные мышцы, как следует из их названия, прикреплены, как правило, к костям скелета; благодаря сокращениям этих мышц поддерживается положение скелета в пространстве и происходят его движения. Сокращения возникают под влиянием импульсов от нервных клеток и обычно бывают произвольными.
На рисунке 4-1 А представлены волокна скелетной мышцы (верхняя панель), клетка сердечной мышцы (средняя панель) и клетка гладкой мышцы (нижняя панель). Клетка скелетной мышцы называется мышечным волокном. В процессе эмбрионального развития каждое мышечное волокно формируется путем слияния многих недифференцированных одноядерных клеток (миобластов) в одну цилиндрическую многоядерную клетку. Дифференцировка
скелетных мышц завершается примерно к моменту рождения. В период от младенческого до взрослого состояния организма размеры дифференцированных мышечных волокон продолжают увеличиваться, но новые волокна из миобластов не образуются. У взрослого человека диаметр мышечных волокон достигает 10-100 мкм, длина - до 20 см.
Если в постнатальный период происходит повреждение скелетных мышечных волокон, они не могут замещаться путем деления сохранившихся волокон, однако новые волокна образуются из недифференцированных клеток, так называемых клеток-сателлитов, расположенных рядом с мышечными волокнами и подвергающихся дифференцировке аналогично эмбриональным миобластам. Возможности формирования новых волокон в скелетной мышце значительны, однако после сильного повреждения она целиком уже не восстанавливается. Важную роль в компенсации
утраченной мышечной ткани играет увеличение неповрежденных мышечных волокон.
На рисунке рис. 4-1 А, Г представлена также сердечная мышца (миокард), которая обеспечивает работу сердца.
Слои гладких мышц находятся в стенках полых внутренних органов и трубчатых образований: желудка, кишечника, мочевого пузыря, матки, кровеносных сосудов, бронхов. В результате сокращений гладких мышц проталкивается содержимое полых органов, регулируется ток жидкости в сосудах и протоках путем изменений их диаметра. Маленькие пучки гладкомышечных клеток находятся также в коже около волосяных сумок и в
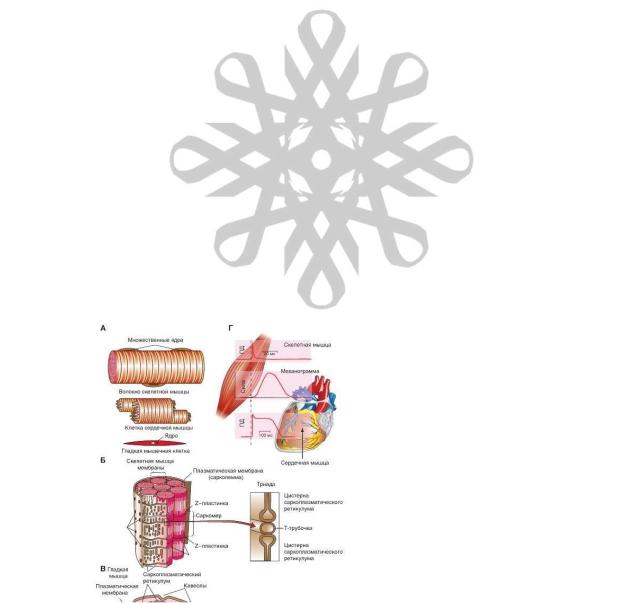
радужной оболочке глаза. Сокращениями гладких мышц управляет вегетативная нервная система, гормоны, аутокринные / паракринные факторы, другие местные химические сигналы. Некоторые гладкие мышцы спонтанно сокращаются даже при отсутствии сигналов. В отличие от скелетных мышц, гладкая мускулатура не имеет произвольной регуляции.
Несмотря на существенные различия между этими тремя видами мышц, у них сходный механизм генерирования силы. Сначала будут рассмотрены скелетные мышцы, затем гладкая мускулатура. Сердечная мышца характеризуется сочетанием ряда свойств двух первых видов мышц.
Наиболее примечательная характеристика волокон скелетной, а также сердечной мышц при изучении с помощью светового микроскопа - чередование светлых и темных полос, поперечных по отношению к длинной оси волокна. Благодаря этой особенности оба типа мышц относят к поперечнополосатым мышцам (рис. 4-1 А, верхняя и средняя панели). В гладкой мышце такая картина отсутствует (рис. 4-1 А, нижняя панель).
В скелетной мышце толстые и тонкие филаменты образуют периодический рисунок вдоль каждой миофибриллы. Регулярно повторяющийся элемент этого рисунка известен как саркомер (от греч. sarco - мышца, mere - маленький) (увеличенный фрагмент на рис. 4-1 Б). Каждый саркомер включает в себя триаду:
1)цистерну саркоплазматического ретикулума;
2)поперечную тубулу;
3)еще одну цистерну саркоплазматического ретикулума (рис. 4-1 Б).
На рисунке 4-1 В представлено строение гладкой мышцы, которое отличается от скелетной.
Совмещенный рисунок 4-1 Г демонстрирует синхронную запись потенциалов действия, а также механограмму скелетной мышцы и мышцы сердца.

Скелетные мышцы
1.Скелетные мышцы состоят из цилиндрических мышечных волокон (клеток); каждый конец мышцы соединен посредством сухожилий с костями.
2.Волокна скелетных мышц характеризуются периодическим чередованием светлых и темных полос, отражающих пространственную организацию толстых и тонких филаментов в миофибриллах.
3.Тонкие филаменты, содержащие актин, прикреплены на обоих краях саркомера к Z- полосам; свободные концы тонких филаментов частично перекрываются с миозинсодержащими толстыми филаментами в А-полосе центральной части саркомера.
4.Во время активного укорочения скелетного мышечного волокна тонкие филаменты подтягиваются по направлению к центру саркомера в результате движений миозиновых поперечных мостиков, которые связываются с актином:
-две глобулярные головки каждого поперечного мостика содержат участок связывания с актином, а также фермент, расщепляющий АТФ;
-каждый рабочий цикл поперечного мостика состоит из четырех стадий. Во время сокращения поперечные мостики совершают повторные циклы, каждый из которых обеспечивает очень маленькое продвижение тонких филаментов;
-АТФ выполняет во время мышечного сокращения три функции.
5.В покоящейся мышце прикрепление поперечных мостиков к актину заблокировано молекулами тропомиозина, контактирующими с субъединицами актина тонких филаментов.
6.Сокращение инициируется в результате повышения цитоплазматической концентрации Са2+. При связывании ионов Са2+ с тропонином изменяется его конформация, благодаря чему тропомиозин смещается, открывая доступ к участкам связывания на молекулах актина; поперечные мостики связываются с тонкими филаментами:
-повышение цитоплазматической концентрации Са2+ запускается потенциалом действия плазматической мембраны. Потенциал действия распространяется вглубь волокна
вдоль поперечных трубочек к саркоплазматическому ретикулуму и вызывает высвобождение Са2+ из ретикулума;
-расслабление мышечного волокна после сокращения происходит в результате активного обратного транспорта Са2+ из цитоплазмы в саркоплазматический ретикулум.
7.Окончания двигательного аксона образуют нервно-мышечные соединения с мышечными волокнами двигательной единицы соответствующего мотонейрона. Каждое мышечное волокно иннервируется ветвью только одного мотонейрона:
-АЦХ, высвобождаемый из двигательных нервных окончаний при поступлении потенциала действия мотонейрона, связывается с рецепторами двигательной концевой пластинки мышечной мембраны; открываются ионные каналы, пропускающие Na+ и К+, благодаря чему концевая пластинка деполяризуется;
-одного потенциала действия мотонейрона достаточно, чтобы вызвать потенциал действия в волокне скелетной мышцы.
8.Существует определенная последовательность процессов, ведущих к сокращению скелетного мышечного волокна.
9.Понятие «сокращение» относится к включению рабочего цикла поперечных мостиков. Изменяется ли при этом длина мышцы, зависит от действия на нее внешних сил.
10.При активации мышечного волокна возможны три типа сокращения:
-изометрическое сокращение, когда мышца генерирует напряжение, но ее длина не меняется;

-изотоническое сокращение, когда мышца укорачивается, перемещая нагрузку;
-удлиняющее сокращение, когда внешняя нагрузка заставляет мышцу удлиняться во время сократительной активности.
11.Повышение частоты потенциалов действия мышечного волокна сопровождается увеличением механической реакции (напряжения или укорочения) до тех пор, пока не будет достигнут максимальный уровень тетанического напряжения.
12.Максимальное изометрическое тетаническое напряжение развивается в случае оптимальной длины саркомера Lо. При растяжении волокна более его оптимальной длины или уменьшении длины волокна менее Lо генерируемое им напряжение падает.
13.Скорость укорочения мышечного волокна снижается при повышении нагрузки. Максимальная скорость соответствует нулевой нагрузке.
14.АТФ образуется в мышечных волокнах следующими способами: перенос фосфата с креатинфосфата к АДФ; окислительное фосфорилирование АДФ в митохондриях; субстратное фосфорилирование АДФ в процессе гликолиза.
15.В начале физического упражнения главным источником энергии служит мышечный гликоген. При более длительном упражнении энергия образуется в основном за счет глюкозы и жирных кислот, поступающих с кровью; по мере дальнейшего продолжения физической деятельности возрастает роль жирных кислот. Когда интенсивность физической работы превысит ~70% от максимума, все более значительную часть образующейся АТФ начинает обеспечивать гликолиз.
16.Мышечное утомление обусловлено рядом факторов, включая изменения кислотности внутриклеточной среды, уменьшение запасов гликогена, нарушение электромеханического сопряжения, но не истощение АТФ.
17.Различают три типа скелетных мышечных волокон в зависимости от максимальной скорости укорочения и преобладающего способа образования АТФ: медленные оксидативные, быстрые оксидативные и быстрые гликолитические:
-разная максимальная скорость укорочения быстрых и медленных волокон обусловлена различиями АТФазы миозина: высокой и низкой АТФазной активности соответствуют быстрые и медленные волокна;
-быстрые гликолитические волокна имеют в среднем больший диаметр, чем оксидативные, и потому развивают более значительное напряжение, однако быстрее утомляются.
18.Все мышечные волокна одной двигательной единицы принадлежат к одному и тому же типу; большинство мышц содержат все три типа двигательных единиц.
19.Известны характеристики трех типов скелетных мышечных волокон.
20.Напряжение целой мышцы зависит от величины напряжения, развиваемого каждым волокном, и от количества активных волокон в мышце.
21.Мышцы, выполняющие тонкие движения, состоят из двигательных единиц с небольшим числом волокон, тогда как большие мышцы, обеспечивающие поддержание позы тела, состоят из гораздо более крупных двигательных единиц.
22.Быстрые гликолитические двигательные единицы содержат волокна большего диаметра и, кроме того, их двигательные единицы имеют более значительное число волокон.
23.Повышение мышечного напряжения происходит прежде всего путем увеличения количества активных двигательных единиц, т.е. их вовлечения. В начале сокращения первыми вовлекаются медленные оксидативные двигательные единицы, затем быстрые оксидативные и, наконец, уже при очень интенсивном сокращении, быстрые гликолитические единицы.
24.Вовлечение двигательных единиц сопровождается повышением скорости, с которой мышца перемещает нагрузку.
25.Силу и утомляемость мышцы можно изменить посредством тренировки:

-продолжительныеупражнения низкой интенсивности повышают способность мышечных волокон к образованию АТФ окислительным (аэробным) путем. Это происходит благодаря увеличению количества митохондрий и кровеносных сосудов в мышце. В итоге возрастает выносливость мышцы;
-кратковременные упражнения высокой интенсивности увеличивают диаметр волокон вследствие повышения синтеза актина и миозина. В итоге возрастает мышечная сила.
26. Движения суставов осуществляются посредством двух антагонистических групп мышц: сгибателей и разгибателей.
27. Мышцы вместе с костями представляют собой системы рычагов; чтобы конечность могла удержать груз, изометрическое напряжение мышцы должно существенно превысить массу этого груза, зато скорость перемещения плечарычага гораздо больше, чем скорость укорочения мышцы.
4.https://helpiks.org/8-34769.html
Одиночное мышечное сокращение - это сокращение мышцы в ответ на раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом
Одиночное мышечное сокращение продолжается около 100 мс и развивается по фазам:
Латентный (скрытый) период продолжается до 3 мс и представляет время от начала
действия раздражителя до начала видимого ответа (сокращения) мышцы.
Фаза сокращения продолжается 40-50 мс
характеризуется укорочением длины мышечного волокна, что связано
с увеличением концентрации Сa2+ в протофибриллярных пространствах и образованием актин-миозиновых связей.
Фаза расслабления продолжается 50-60 мс характеризуется увеличением (восстановлением) длины волокна. Возникает при
снижении концентрации Ca2+ в протофибриллярных пространствах и ослаблением актин-миозиновых связей.
Если на мышцу наносятся два и более раздражений с интервалом менее
продолжительности одиночного сокращения, но более продолжительности рефрактрного периода ПД, то
происходит суммация сокращений, в результате которой сократительный эффект усиливается.
Существует два типа суммации: частичная и полная
Частичная (или неполная) суммация возникает, если
·интервал между раздражениями меньше продолжительности одиночного мышечного сокращения;
·больше продолжительности фазы сокращения, т.е. если второе
раздражение попадает в фазу расслабления.
В результате амплитуда мышечного сокращения возрастает с образованием двух вершин.
Полная суммация возникает, если:
· интервал между раздражениями меньше продолжительности фазы сокращения, но больше продолжительности рефрактерного периода;

·второе раздражение попадает в фазу сокращения.
Врезультате амплитуда мышечного сокращения изменяется (увеличивается или уменьшается относительно одиночного сокращения) с
образованием одной вершины
Увеличение или уменьшение амплитуды связано с изменением возбудимости в процессе возбуждения и зависит от того, в какую фазу измененной возбудимости наносится следующее раздражение.
Учитывая, что в скелетной мышце процесс возбуждения продолжается около 8 мс (латентный период ПД - 2,5 мс плюс пиковый потенциал – около 5 мс), становится понятным, что укорочение мышечного волокна начнется тогда, когда быстрая деполяризация произойдет приблизительно на 1/3 от амплитуды пикового потенциала. Известно, что в период формирования пикового потенциала возбудимость ткани снижена (фаза абсолютной и фаза относительной рефрактерности). Поэтому, если следующее раздражение будет наноситься в этот период, то амплитуда мышечного сокращения будет снижена.
Период возбуждения в скелетной мышце завершается следовой деполяризацией, продолжающейся от 20 до 40 мс.
В этот период возбудимость, а, следовательно, и сократимость повышена. Поэтому, если следующее раздражение будет приходиться на этот период, то амплитуда мышечного сокращения будет возрастать (тем больше, чем больше повышена возбудимость).
5. https://studopedia.ru/19_315059_vopros--odinochnie-sokrashcheniya-i-ego-vidi-tetanus- faktori- vilyayushchie-na-ego-velichinu.html
Тетанусы: в физиологии, длительное сокращение мышц, возникающее при последовательном воздействии на них ряда нервных импульсов, разделённых малыми интервалами, и основанное на временной суммации следующих друг за другом одиночных волн сокращения. Т. наступает при достаточно высокой частоте возбуждения мышцы, когда каждое новое сокращение возникает до окончания предыдущего; при этом сократительные волны как бы накладываются друг на друга (слитное сокращение); в результате мышца остаётся укороченной в течение всего периода раздражения.
Различают зубчатый и гладкий Т.
Зубчатый Т. наблюдается в тех случаях, когда в ответ на последующее (второе, третье и т. д.) раздражение мышца начинает сокращаться, не успев полностью расслабиться после предыдущего сокращения.
Гладкий Т. образуется при более высокой частоте раздражения, когда каждый последующий стимул приходит в фазу укорочения мышцы. Суммационная природа Т. подтверждается тем, что во время Т. в мышце ритмически возникают электрические потенциалы действия, сопровождающие каждую вспышку возбуждения. Тетаническое сокращение по амплитуде и длительности
значительно превосходит одиночное сокращение. Характер Т. определяется тем, в какую фазу возбудимости мышцы (например, фазы экзальтации, рефрактерности) приходит очередное раздражение. На зависимость величины Т.

от уровня возбудимости мышцы впервые указал Н. Е. Введенский, который отметил, что при повышении частоты раздражения Т. вначале достигает максимальной амплитуды (Оптимум), а затем амплитуда Т. резко снижается (Пессимум). Для тетанически сократившихся мышечных волокон характерна относительно быстрая утомляемость, так как Т. сопровождается значительным расходованием энергетических ресурсов мышцы.
Пессимум (от лат. pessimum — наихудшее) -(физиологическое), угнетение деятельности органа или ткани, вызываемое чрезмерной частотой или силой наносимых раздражений; описано в 1886 Н. Е. Введенским (См. Введенский).
Исследуя особенности проведения нервного импульса в нервно-мышечном препарате лягушки, он обнаружил, что усиление слитного сокращения мышцы — так называемое Тетануса, вызываемое постепенным возрастанием частоты или силы раздражений (см. Оптимум), при дальнейшем их учащении или усилении, внезапно сменяется расслаблением мышцы и полным торможением её активности. Введенский трактовал это явление с позиций разработанной им теории Парабиоза. Согласно этой теории, работоспособность нервных окончаний,
передающих импульсы мышце, после прохождения волны возбуждения резко падает, и для восстановления их работоспособности требуется некоторое время (в нервно-мышечном препарате икроножной мышцы лягушки — 0,02—0,03 сек). Это время определяет функциональные возможности нервных окончаний — их Лабильность. Если интервал между раздражениями меньше этого необходимого
периода, то есть если он превышает лабильность нервных окончаний, в них развивается своеобразное стойкое нераспространяющееся возбуждение — парабиоз, блокирующее проведение нервных импульсов к мышце и тормозящее тем
самым её активность, предохраняя от переутомления. Описываемое явление носит обратимый характер: снижение интенсивности раздражения восстанавливает мышечное сокращение. Явление П. обнаружено в ряде органов и тканей;
многие исследователи полагают, что оно лежит в основе рефлекторной регуляции деятельности организма со стороны нервной системы.
Оптимум (от лат. optimum — наилучшее) - уровень силы или частоты
раздражений, при котором осуществляется максимальная деятельность органа или ткани. Явление О. описано в 1886 Н. Е. Введенским (См. Введенский), который на нервномышечном препарате лягушки установил, что нарастание до некоторого предела частоты или силы раздражений усиливает длительное, слитное сокращение мышцы — Тетанус. О. объясняют тем, что в этих случаях каждое последующее раздражение падает на мышцу в период повышенной её возбудимости, вызванной предыдущим раздражением.
6. https://studopedia.ru/8_109636_sila-rabota-i-utomlenie-mishts.html
Сила - мера механического воздействия на мышцу со стороны других тел, которая выражается в ньютонах или кг-силах. При изотоническом сокращении в
эксперименте сила определяется массой максимального груза, который мышца может поднять (динамическая сила), при изометрическом - максимальным напряжением, которое она может развить (статическая сила).
В различных мышцах тела соотношение между числом медлен-,, ных и быстрых мышечных волокон неодинаково, поэтому и сила их сокращения, и степень укорочения вариабельны.

При снижении физической нагрузки - особенно большой интенсивности, при которой требуется активное участие быстрых мышечных волокон, - последние истончаются (гипотрофируются) быстрее, чем медленные волокна, быстрее уменьшается их число.
А. Факторы, влияющие на силу сокращения мышцы.
1.Число сокращающихся волокон в данной мышце. С увеличением сокращающихся волокон возрастает сила сокращений мышцы в целом. В
естественных условиях сила сокращения мышцы возрастает с увеличением нервных импульсов, поступающих к мышце, в эксперименте - с увеличением силы раздражения.
2.Соотношение быстрых и медленных волокон. Чем больше быстрых волокон содержит мышца, тем больше возможная ее сила сокращения.
3.Поперечное сечение мышцы. Различают геометрическое и
физиологическое поперечные сечения мышцы. Геометрическое поперечное сечение перпендикулярно продольной оси мышцы, физиологическое - длине мышечных волокон. В портняжной мышце, например, все волокна параллельны длине оси мышцы - парал-
лельноволокнистый тип. Большинство мышц – перистого типа, их волокна расположены косо,
прикрепляясь с одной стороны к центральному сухожильному тяжу, а с другой стороны - к наружному сухожильному футляру. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон физиологическое поперечное сечение может значительно превосходить геометрическое поперечное сечение.
7. https://studopedia.ru/8_109636_sila-rabota-i-utomlenie-mishts.html
Утомление - временное снижение или потеря работоспособности отдельной клетки, ткани, органа или организма в целом, наступающее после нагрузок (деятельности). Утомление мышц происходит при их длительном сокращении (работе) и имеет определенное биологическое значение, сигнализируя о истощении (частичном) энергетических ресурсов.
При утомлении понижаются функциональные свойства мышцы: возбудимость, лабильность и сократимость. Высота сокращения мышцы при развитии утомления постепенно снижается. Это снижение может дойти до полного исчезновения сокращений. Понижаясь, сокращения делаются все более растянутыми, особенно за счет удлинения периода расслабления: по окончании сокращения мышца долго не возвращается к первоначальной длине, находясь в состоянии контрактуры (крайне замедленное расслабление мышцы). Скелетные мышцы утомляются раньше гладких. В скелетных мышцах сначала утомляются белые волокна, а потом красные.
Из различных представлений о механизме утомления одной из наиболее ранних теорий, объясняющих утомление, была теория истощения, предложенная К. Шиффом. Согласно этой теории причиной утомления
служит исчезновение в мышце энергетических веществ, в частности гликогена. Однако, детальное изучение показало, что в

утомленных до предела мышцах содержание гликогена еще значительно. В дальнейшем Е. Пфлюгером была выдвинута теория засорения органа продуктами рабочего распада (теория отравления). Согласно этой теории, утомление объясняется накоплением большого
количества молочной, фосфорной кислот и недостатком кислорода, а так же других продуктов обмена, которые нарушают обмен веществ в работающем органе и его деятельность прекращается.
Обе эти теории сформулированы на основании данных, полученных в экспериментах на изолированной скелетной мышце и объясняют утомление односторонне и упрощенно. Дальнейшим изучением утомления в условиях целого организма установлено, что в утомленной мышце появляются продукты обмена веществ, уменьшается содержание гликогена, АТФ, креатинофосфата. Изменения наступают в сократительных белках мышцы. Происходит связывание или уменьшение сульфгидрильных групп актомиозина, в результате чего нарушается процесс синтеза и распада АТФ. Нарушения в химическом составе мышцы, находящейся в целостном организме, выражены в меньшей степени, чем в изолированной благодаря транспортной функции крови.
Исследованиями Н.Е. Введенского установлено, что утомление прежде всего развивается в нервно-мышечном синапсе в связи с низкой его лабильностью. Быстрая утомляемость синапсов обусловлена несколькими факторами.
Во-первых, при длительном раздражении в нервных окончаниях уменьшается запас медиатора, а его синтез не поспевает за расходованием. Во-вторых, накапливающиеся продукты обмена в мышце понижают чувствительность постсинаптической мембраны к ацетилхолину, в результате чего уменьшается величина постсинаптического потенциала.
Когда он понижается до критического уровня, в мышечном волокне не возникает возбуждения.
И.М.Сеченов (1903), исследуя на сконструированном им эргографе для двух рук работоспособность мышц при поднятии груза, установил, что работоспособность утомленной правой руки восстанавливается полнее и быстрее после активного отдыха, т.е. отдыха сопровождаемого работой левой руки. Подобного же рода влияние на работоспособность утомленной руки оказывает сочетающееся с отдыхом раздражение индукционным током чувствительных (афферентных) нервных волокон кисти другой руки, а также работа ногами, связанная с подъемом тяжести, и вообще двигательная активность.
Таким образом, активный отдых, сопровождающийся умеренной работой других мышечных групп, оказывается более эффективным средством борьбы с утомлением двигательного аппарата, чем простой покой.
Причину наиболее эффективного восстановления работоспособности двигательного аппарата в условиях активного отдыха Сеченов с полным основанием связывал с действием на центральную нервную систему